Photo credit: Google
Fruit of Knema globularia
Comparative leaf anatomy of the Asiatic Myristicaceae
by Koster J., Baas P. (1981)
Rijksherbarium Leiden, The Netherlands
in Blumea 27: 115–173 –
Google Scholar –
http://repository.naturalis.nl/document/565506
The stomatal complex (photos 13, 23—27)
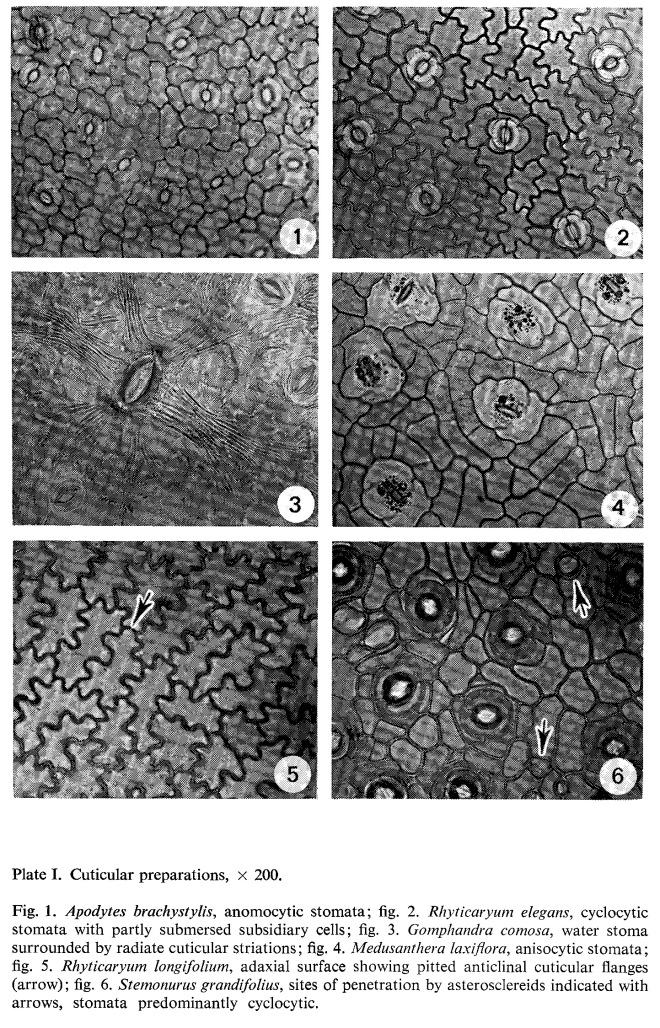
Stomata are confined to the abaxial epidermis; however, in Gymnacranthera bancana var. borneensis a very small number is also present in the adaxial epidermis. They are randomly distributed, abundant on the areolae, scanty on the midrib, the veins and along the leaf margin. If they are present here, the stomatal complex shows abnormalities.
Several authors have pointed out that the stomatal index may be extremely variable within a species (Bongers, 1973; Jansen & Baas, 1973; Van Staveren & Baas, 1973). Stomatal indices have therefore not been determined in this study.
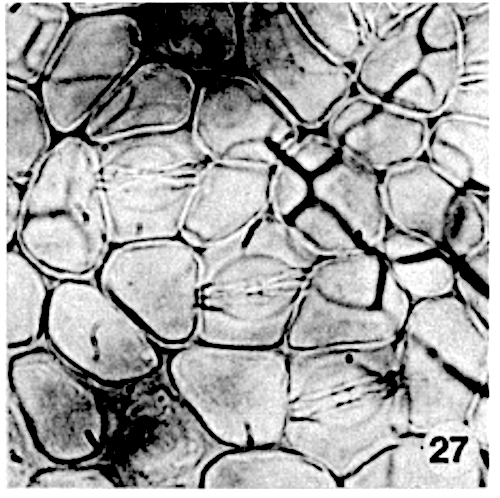
The stomatal type of the species examined is paracytic (photo 27). The dimensions of the guard cell pairs range from 8 to 21 mu for the width and from 15 to 39 mu for the length. The genera have a different size range (in Myristica relatively long stomata and in Gymnacranthera and Knema relatively short stomata are present), but a large overlap occurs.
The variation in one single leaf can be up to 8 and 12 mu for the width and length respectively. However, the average values show fairly little variation in a genus. Therefore, the data for the stomatal sizes per specimen are not given in tables.
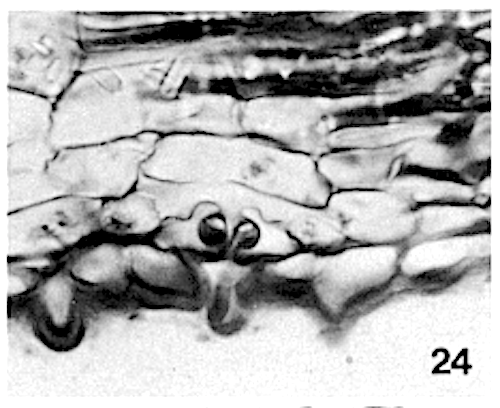
Giant or water stomata have been recorded for some specimens of Gymnacranthera contracta and Gymnacranthera forbesii var. forbesii only. The guard cells are often embedded in the subsidiary cells, most conspicuously in Gymnacranthera and Knema; here, in a section perpendicular to the pore, the subsidiary cells fully enclose the guard cells, the poral sides excepted (photo 24).
In Gymnacranthera such sections also show pointed, sometimes protruding subsidiary cells, whereas the unspecialized epidermal cells are lowly dome-shaped or square to rectangular. Outer stomatal ledges are fairly well developed in some species of Horsfieldia only.
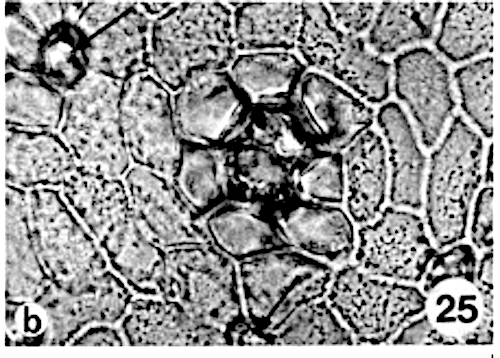
A peculiar character is formed by the cells surrounding the stomatal complex in Knema and Myristica. In the cuticular macerations and the free hand sections of the Knema species the stomatal complex itself is difficult to observe, lying on a lower level than the other epidermal cells.
A star-shaped opening, left by 4 to 6 epidermal cells overarching the stomatal complex, indicates the position of the stoma (photo 23). The transverse sections show the strongly sunken stomatal complex with a thin cuticle, overarched by more or less horizontally directed papillae of the bordering epidermal cells (photo 24); these nipple-shaped papillae give the star its characteristic form.
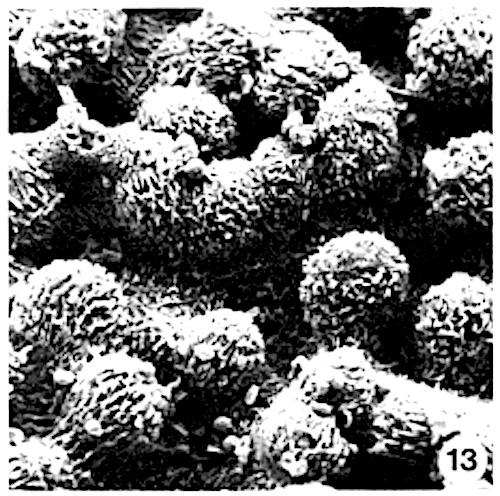
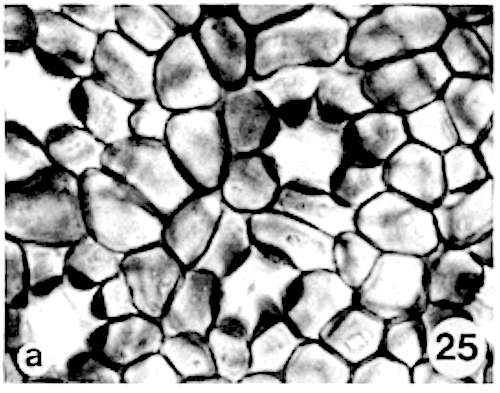
Siddiqi and Wilson (1975c) proposed that the ‘star-shaped structure’ is formed by hypodermal cells. This appears to be incorrect. In Myristica the sunken stomatal complex is partly overarched by more or less upright papillae of the 4 to 11 bordering epidermal cells (photo 26). In surface view these papillae, varying in height per species, form a ring above the stomatal complex (photo 13,25). As stated elsewhere, the unspecialized epidermal cells in most species are without papillae.







{kind=link}
You must be logged in to post a comment.