
Parasitism of Corn Earworm, Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae), by Tachinid Flies in Cultivated Hemp

 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Field Work
2.2. Laboratory Measurements
2.3. Data Analysis
2.4. Mortality of CEW and Tachinid Emergence
3. Results
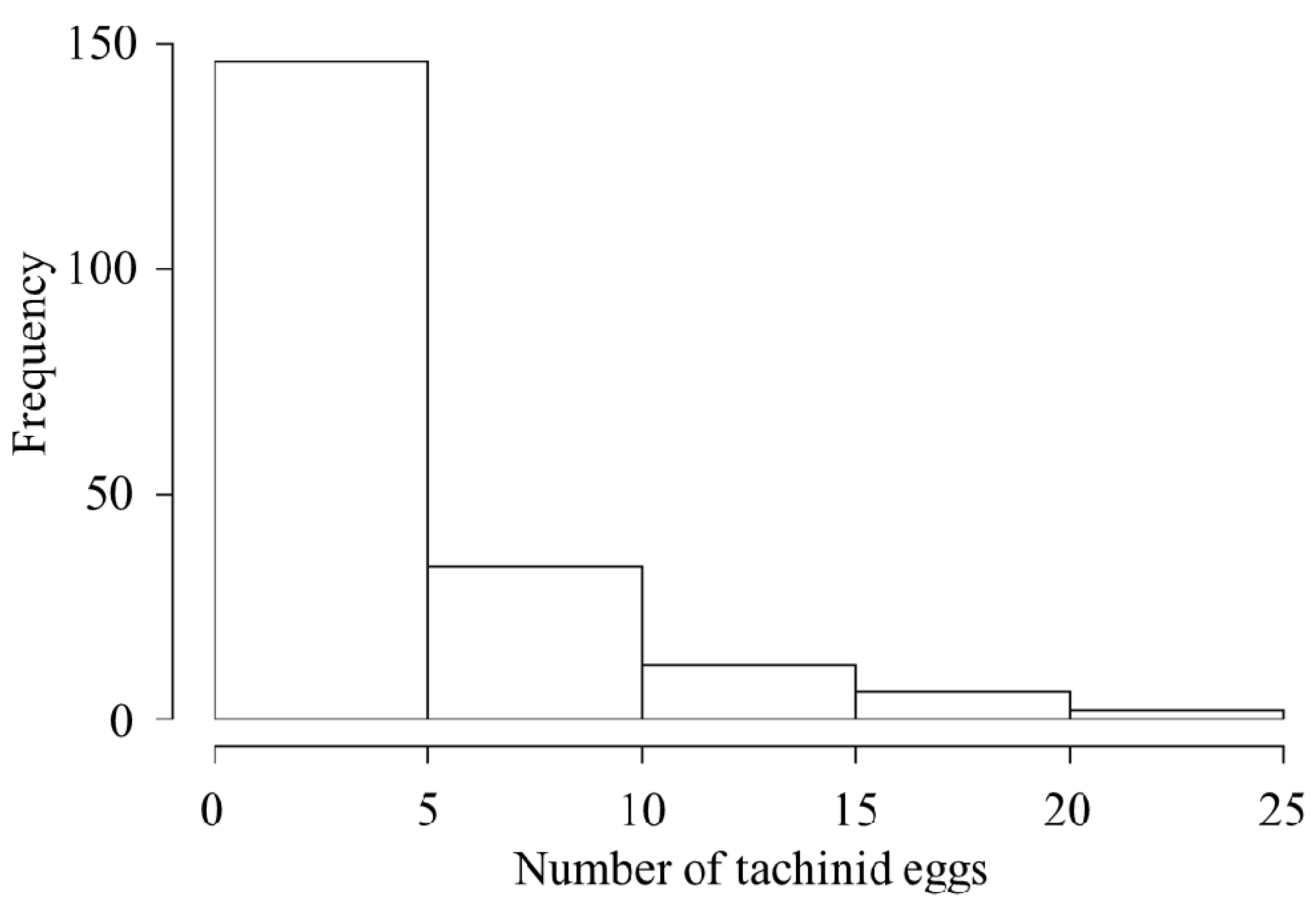
3.1. Parasitism Frequency
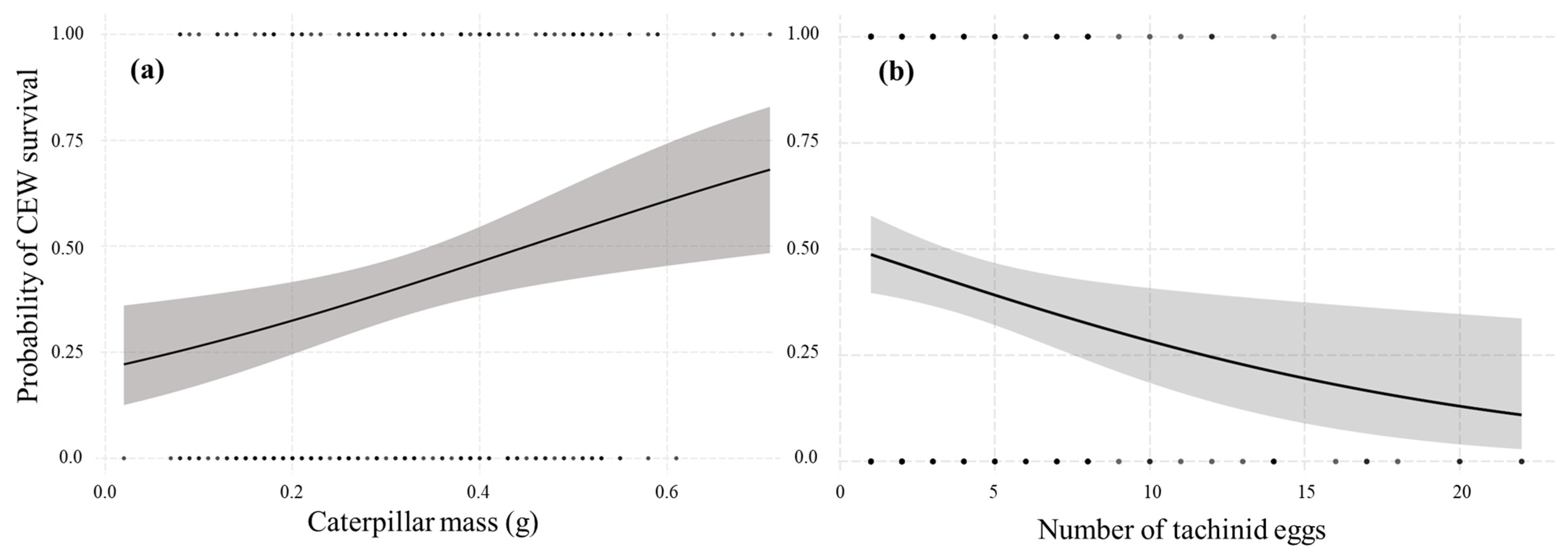
3.2. Mortality
4. Discussion
4.1. Parasitism
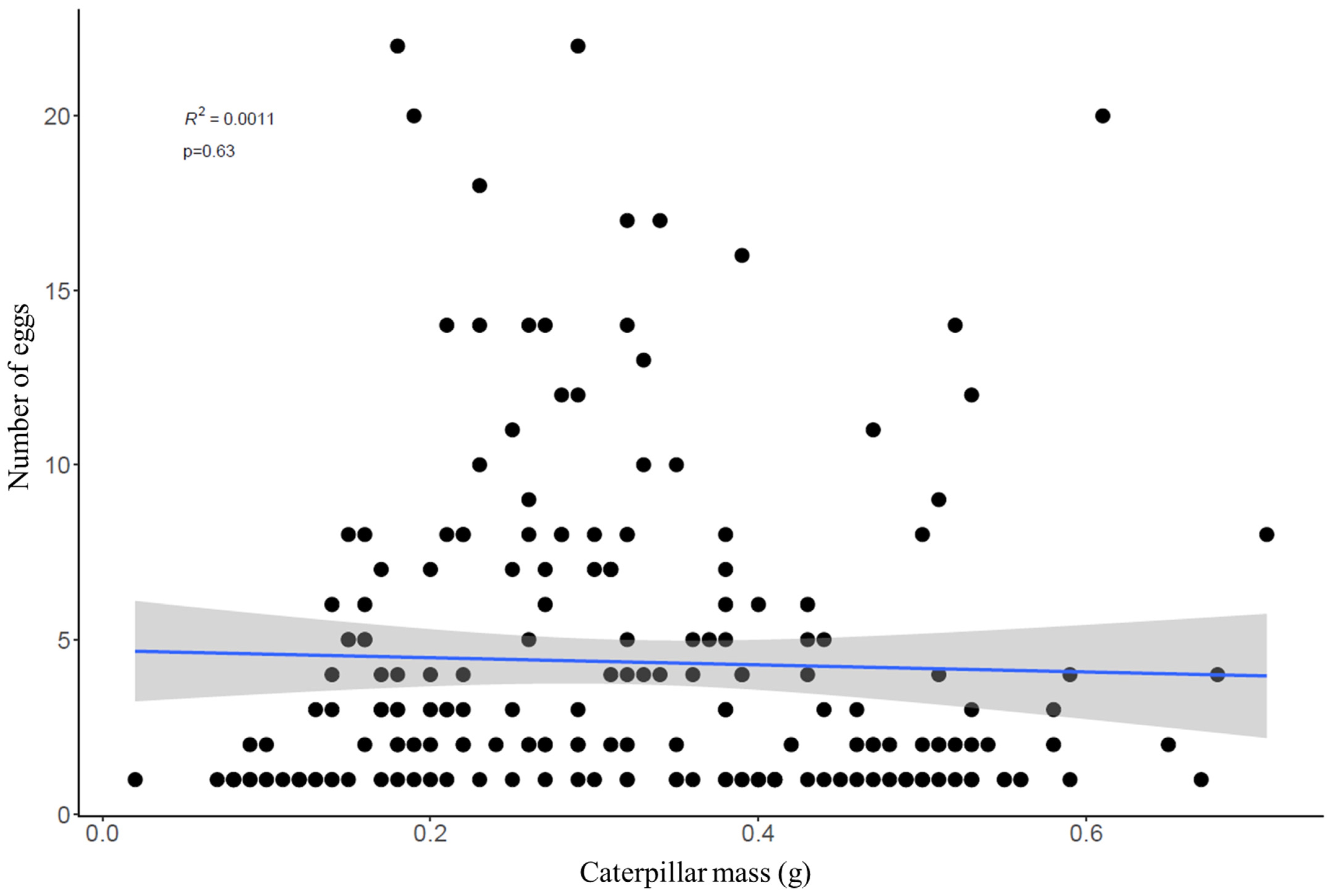
4.2. Parasitoid Egg Numbers and Distribution
4.3. Host Mortality and Parasitoid Success
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deitch, R. Hemp: American History Revisited: The Plant with a Divided History; Algora Publishing: New York, NY, USA, 2003. [Google Scholar]
- Cranshaw, W.; Schreiner, M.; Britt, K.; Huhar, T.P.; McPartland, J.; Grant, J. Developing Insect Pest Management Systems for Hemp in the United States: A Work in Progress. J. Integr. Pest Manag. 2019, 10, 26. [Google Scholar] [CrossRef]
- Agricultural Marketing Service, USDA. Hemp Production. Available online: https://www.ams.usda.gov/rules-regulations/hemp (accessed on 2 February 2022).
- Salentijn, E.M.J.; Zhang, Q.; Amaducci, S.; Yang, M.; Trindade, L.M. New developments in fiber hemp (Cannabis sativa L.) breeding. Ind. Crop. Prod. 2015, 68, 32–41. [Google Scholar] [CrossRef]
- Mark, T.; Shepherd, J. Trojan Horse or Golden Ticket. In Science of Hemp: Production and Pest Management; University of Kentucky, College of Agriculture, Food and Environment: Lexington, KY, USA, 2020; p. 33. [Google Scholar]
- McPartland, J.M.; Clarke, R.C.; Watson, D.P. Hemp Diseases and Pests, Management and Biological Control; CABI: New York, NY, USA, 2000. [Google Scholar]
- Batra, S.W.T. Some insects associated with hemp or marijuana (Cannabis sativa L.) in northern India. J. Kans. Entomol. Soc. 1976, 49, 385–388. [Google Scholar]
- Lago, P.K.; Stanford, D.F. Phytophagous insects associated with cultivated marijuana, Cannabis sativa, in northern Mississippi. Entomol. Sci. 1989, 24, 437–445. [Google Scholar] [CrossRef]
- McPartland, J.M. Cannabis pests. JIHA. 1996, 3, 52–55. [Google Scholar]
- Edde, P.A. Field Crop Arthropod Pests of Economic Importance; Elsevier; AP Academic Press: San Diego, CA, USA, 2021. [Google Scholar]
- Ajayi, O.S.; Samuel-Foo, M. Hemp pest spectrum and potential relationship between Helicoverpa zea infestation and hemp production in the United States in the face of climate change. Insects 2021, 12, 940. [Google Scholar] [CrossRef]
- Villanueva, R.T. An Update in hemp insects: Emphasizing key pests in Kentucky in 2019. In Science of Hemp: Production and Pest Management; University of Kentucky, College of Agriculture, Food and Environment: Lexington, KY, USA, 2020; p. 48. [Google Scholar]
- EPA (United States Environmental Protection Agency). Pesticide Products Registered for Use on Hemp. 2022. Available online: https://www.epa.gov/pesticide-registration/pesticide-products-registered-use-hemp (accessed on 15 February 2022).
- Grant, J.; Hale, F. Beneficials on Hemp: What You Need to Know. In Proceedings of the Entomological Society of America Annual Meeting, Virtual, 16–19 November 2020; pp. 15–18. [Google Scholar]
- Bryan, D.E.; Jackson, C.G.; Patana, R. Laboratory Studies of Lespesia archippivora in Four Lepidopterous Hosts. J. Econ. Entomol. 1968, 61, 819–823. [Google Scholar] [CrossRef]
- Danks, H.V. Seasonal cycle and biology of Winthemia rufopicta (Diptera: Tachinidae) as a parasite of Heliotis spp. (Lepidoptera: Noctuidae) on tobacco in North Carolina. Can. Entomol. 1975, 107, 639–654. [Google Scholar] [CrossRef]
- Danks, H.V. Factors determining levels of parasitism by Winthemia rufopicta (Diptera: Tachinidae), with particular reference to Heliothis spp. (Lepidoptera: Noctuidae) as hosts. Can. Entomol. 1975, 107, 655–684. [Google Scholar] [CrossRef]
- Proshold, F.I.; Gross, H.R.; Carpenter, J.E. Inundative release of Archytas marmoratus (Diptera: Tachinidae) against the corn earworm and fall armyworm (Lepidoptera: Noctuidae) in whorl-stage corn. Entomol. Sci. 1988, 33, 241–255. [Google Scholar] [CrossRef]
- Barbosa, P.; Caldas, A. Do larvae of species in macrolepidopteran assemblages share traits that influence susceptibility to parasitism? Environ. Entomol. 2007, 36, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Carper, A.L.; Enger, A.; Bowers, M.D. Host plant effects on immune response across development of a specialist caterpillar. Front. Ecol. Evol. 2019, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, J.E.; Proshold, F.I. Survival of Archytas marmoratus (Diptera: Tachinidae) from superparasitized corn earworm larvae (Lepidoptera: Noctuidae). Environ. Entomol. 2000, 29, 606–611. [Google Scholar] [CrossRef]
- Guimarães, J.H. A revision of the genus Winthemia Robineau-Desvoidy in America north of Mexico (Diptera, Tachinidae). Arq. Zool. 1972, 22, 27–112. [Google Scholar] [CrossRef] [Green Version]
- Sabrosky, C.W. Identification of Winthemia of America north of Mexico, with a revised key to the females (Diptera, Tachinidae). Ann. Entomol. Soc. Am. 1973, 66, 1035–1041. [Google Scholar] [CrossRef]
- Sabrosky, C.W. A revised key to the Nearctic species of Lespesia (Diptera: Tachinidae). Ann. Entomol. Soc. Am. 1980, 73, 63–73. [Google Scholar] [CrossRef]
- Danks, H.V.; Rabb, R.L.; Southern, P.S. Biology of insect parasites of Heliothis larvae in North Carolina. J. Georgia Entomol. Soc. 1979, 14, 36–64. [Google Scholar]
- Hardwick, D.F. The corn earworm complex. Mem. Ent. Soc. Can. 1965, 97, 5–247. [Google Scholar] [CrossRef]
- Reitz, S. Superparasitism and intraspecific competition by the solitary larval-pupal parasitoid Archytas marmoratus (Diptera: Tachinidae). Fla. Entomol. 1995, 78, 578–585. [Google Scholar] [CrossRef]
- Henderson, C.F.; Tilton, E.W. Tests with Acaricides against the Brown Wheat Mite. J. Econ. Entomol. 1955, 48, 157–161. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 14 October 2021).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in R. R package version 3.2-13. 2021. Available online: https://CRAN.R-project.org/package=survival (accessed on 8 November 2021).
- Benneway, D.F. A revision of the flies of the genus Lespesia (=Achaetoneura) in North America (Diptera: Tachinidae). Univ. Kansas Sci. Bull. 1963, 44, 627–686. [Google Scholar]
- Arnaud, P.H., Jr. A Host-Parasite Catalog of North American Tachinidae (Diptera); United States Department of Agriculture: Washington, DC, USA, 1978; Volume 1319. [Google Scholar]
- Stireman, J.O., III; Singer, M.S. What determines host range in parasitoids? An analysis of a tachinid parasitoid community. Oecologia 2003, 135, 629–638. [Google Scholar]
- Smith, M.A.; Wood, D.M.; Janzen, D.H.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes affirm that 16 species of apparently generalist tropical parasitoid flies (Diptera, Tachinidae) are not all generalists. Proc. Natl. Acad. Sci. USA 2007, 104, 4967–4972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoza, Y.J.; Epsky, N.D.; Heath, R.R. Biology and development of Lespesia aletiae (Diptera: Tachinidae) in two lepidopteran species in the laboratory. Fla. Entomol. 1997, 80, 289–300. [Google Scholar] [CrossRef]
- Sherman, F. Rearing of moths and tachina-flies from larvae and pupae of army-worm in North Carolina in 1914. J. Econ. Entomol. 1915, 8, 299–302. [Google Scholar] [CrossRef]
- Chauthani, A.R.; Hamm, J.J. Biology of the exotic parasite Drino munda (Diptera: Tachinidae). Ann. Ent. Soc. Am. 1967, 60, 373–376. [Google Scholar] [CrossRef]
- Rahman, M. Exorista flaviceps, a tachinid parasite of Pieris rapae in South Australia. J. Econ. Ent. 1970, 63, 836–841. [Google Scholar] [CrossRef]
- Stireman, J.O., III; O’Hara, J.E.; Wood, D.M. TACHINIDAE: Evolution, Behavior, and Ecology. Annu. Rev. Entomol. 2006, 51, 525–555. [Google Scholar]
- Dindo, M.L.; Nakamura, S. Oviposition strategies of tachinid parasitoids: Two Exorista species as case studies. Int. J. Insect Sci. 2018, 10, 1179543318757491. [Google Scholar] [CrossRef] [Green Version]
- Stireman, J.O., III. Host location and acceptance in a polyphagous tachinid parasitoid. Entomol. Exp. Appl. 2002, 103, 23–34. [Google Scholar] [CrossRef]
- Ichiki, R.T.; Kainoh, Y.; Kugimiya, S.; Yamawaki, Y.; Nakamura, S. The parasitoid fly Exorista japonica uses visual and olfactory cues to locate herbivore-infested plants. Ent. Exp. Appl. 2011, 138, 175–183. [Google Scholar] [CrossRef]
- Nakamura, S. Ovipositional behaviour of the parasitoid fly, Exorista japonica (Diptera: Tachinidae), in the laboratory: Diel periodicity and egg distribution on a host. Appl. Entomol. Zool. 1997, 32, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Morewood, D.W.; Wood, M.D. Host utilization by Exorista thula Wood (sp. nov.) and Chetogena gelida (Coquillett) (Diptera: Tachinidae), parasitoids of arctic Gynaephora species (Lepidoptera: Lymantriidae). Polar Biol. 2002, 25, 575–582. [Google Scholar] [CrossRef]
- Lill, J.T.; Marquis, R.J.; Ricklefs, R.E. Host plants influence parasitism of forest caterpillars. Nature 2002, 417, 170–173. [Google Scholar] [CrossRef]
- Stireman, J.O., III; Singer, M.S. Determinants of parasitoid-host associations: Insights from a natural Tachinid–Lepidopteran community. Ecology 2003, 84, 296–310. [Google Scholar] [CrossRef]
- Stireman, J.O., III; Dyer, L.A.; Greeney, H.F. Specialised generalists? Food web structure of a tropical tachinid-caterpillar community. Insect Conserv. Divers. 2017, 10, 367–384. [Google Scholar] [CrossRef]
- Gerard, P.J.; Charles, J.G.; McNeill, M.R.; Hardwick, S.; Malipatil, M.B.; Page, F.D. Parasitoids of the painted apple moth Teia anartoides Walker (Lepidoptera: Lymantriidae) in Australia. Aust. J. Entomol. 2011, 50, 281–289. [Google Scholar] [CrossRef]
- Badillo-Montaño, R.; Amancio, G.; Falcon-Brindis, A.; León-Cortés, J.; Von-Thaden, J.; Dzul-Cauich, F. Trophic host-parasitoid interactions of two Neotropical butterfly species in southeastern Mexico. Int. J. Trop. Insect Sci. 2022, 42, 1865–1875. [Google Scholar] [CrossRef]
- Morrison, G.; Lewis, W.J.; Nordlund, D.A. Spatial differences in Heliothis zea egg density and the intensity of parasitism by Trichogramma spp.: An experimental analysis. Environ. Entomol. 1980, 9, 79–85. [Google Scholar] [CrossRef]
- Tillman, G.P.; Laster, M.L.; Powell, J.E. Development of the Endoparasitoids Microplitis croceipes, Microplitis demolitor, Cotesia kazak (Hymenoptera: Braconidae) on Helicoverpa zea and H. armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 1993, 86, 360–362. [Google Scholar] [CrossRef]
- Tipping, P.W.; Holko, C.A.; Bean, R.A. Helicoverpa zea (Lepidoptera: Noctuidae) dynamics and parasitism in Maryland soybean. Fla. Entomol. 2005, 88, 55–60. [Google Scholar] [CrossRef]
- Stireman, J.O., III; Shaw, S.R. Natural History and Ecology of Caterpillar Parasitoids. In Caterpillar in the Middle: Tritrophic Interactions in a Changing World; Marquis, R.J., Koptur, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 225–272. [Google Scholar]
- De Morales, C.M.; Lewis, W.J.; Tumlison, J.H. Examining plant-parasitoid interactions in tritrophic systems. An. Soc. Entomol. Bras. 2000, 29, 189–203. [Google Scholar] [CrossRef]
- Smilanich, A.M.; Dyer, L.A.; Chambers, J.Q.; Bowers, M.D. Immunological cost of chemical defence and the evolution of herbivore diet breadth. Ecol. Lett. 2009, 12, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Ode, P.J. Plant chemistry and natural enemy fitness: Effects on Herbivore and Natural Enemy Interactions. Annu. Rev. Entomol. 2006, 51, 163–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slinn, H.L.; Richards, L.A.; Dyer, L.A.; Hurtado, P.J.; Smilanich, A.M. Across multiple species, phytochemical diversity and herbivore diet breadth have cascading effects on herbivore immunity and parasitism in a tropical model system. Front. Plant Sci. 2018, 9, 656. [Google Scholar] [CrossRef]
- Liu, Z.; Li, D.; Gong, P.; Wu, K. Life table studies of the Cotton Bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae), on different host plants. Environ. Entomol. 2004, 33, 1570–1576. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Mean | ±SE | Range (Min–Max) |
|---|---|---|---|
| Body mass | 0.32 g | 0.01 | 0.02–0.71 g |
| Tachinid eggs/host | 4.36 | 0.31 | 1–22 |
| No. of tachinid eggs on thorax | 4.03 | 0.29 | 0–20 |
| No. of tachinid eggs on middle | 0.33 | 0.06 | 0–5 |
| No. of tachinid eggs on posterior | 0.12 | 0.03 | 0–3 |
| Date | N | Mortality (%) | Tachinid Species |
|---|---|---|---|
| 14 September 2021 | 46 | 33 | W. rufopicta (52%), L. aletiae (48%) |
| 17 September 2021 | 11 | 82 | W. rufopicta |
| 20 September 2021 | 38 | 71 | W. rufopicta |
| 23 September 2021 | 20 | 75 | W. rufopicta |
| 27 September 2021 | 24 | 83 | W. rufopicta (57%), L. aletiae (43%) |
| 01 October 2021 | 6 | 17 | L. aletiae |
| 20 October 2021 | 55 | 85 | W. rufopicta (14%), L. aletiae (86%) |
| Survival/Success | Variable | Estimate | SE | p-Value | OR |
|---|---|---|---|---|---|
| CEW | Intercept | −0.89 | 0.392 | 0.023 * | 0.41 |
| CEW body mass | 2.91 | 1.013 | 0.003 ** | 18.52 | |
| Number of tachinid eggs | −0.09 | 0.038 | 0.011 * | 0.91 | |
| Adult flies | Intercept | −3.21 | 0.748 | <0.001 *** | 0.04 |
| CEW body mass | −0.52 | 1.985 | 0.793 | 0.59 | |
| Number of tachinid eggs | 0.16 | 0.043 | <0.001 *** | 1.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falcon-Brindis, A.; Stireman, J.O., III; Viloria, Z.J.; Villanueva, R.T. Parasitism of Corn Earworm, Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae), by Tachinid Flies in Cultivated Hemp. Insects 2022, 13, 519. https://doi.org/10.3390/insects13060519
Falcon-Brindis A, Stireman JO III, Viloria ZJ, Villanueva RT. Parasitism of Corn Earworm, Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae), by Tachinid Flies in Cultivated Hemp. Insects. 2022; 13(6):519. https://doi.org/10.3390/insects13060519
Chicago/Turabian StyleFalcon-Brindis, Armando, John O. Stireman, III, Zenaida J. Viloria, and Raul T. Villanueva. 2022. "Parasitism of Corn Earworm, Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae), by Tachinid Flies in Cultivated Hemp" Insects 13, no. 6: 519. https://doi.org/10.3390/insects13060519