
Mitigating the Mistletoe Menace: Biotechnological and Smart Management Approaches
by
 , and
, and
Gaurav Mudgal
1,*,
Jaspreet Kaur
1,
Kartar Chand
1,
Manisha Parashar
1,
Sanjoy K. Dhar
1,
Gajendra B. Singh
1 and
and
Mayank A. Gururani
2,* 1
University Institute of Biotechnology, Chandigarh University, Mohali 140413, Punjab, India
2
Department of Biology, College of Science, United Arab Emirates University, Al Ain 15551, United Arab Emirates
*
Authors to whom correspondence should be addressed.
Biology 2022, 11(11), 1645; https://doi.org/10.3390/biology11111645
Submission received: 9 October 2022
/
Revised: 8 November 2022
/
Accepted: 8 November 2022
/
Published: 10 November 2022
(This article belongs to the Section Plant Science)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
In this article, we discuss the current status of conventional management and control approaches for mistletoes, the hemiparasitic plants that have emerged as serious pests of trees in forests and commercial fruit and timber plantations. We highlight the research and implementation gaps and discuss possible newer biotechnological interventions to be made in relation to biological control agents and mistletoe-resistant tree cultivars. We also discuss the potential of smart technological alternatives that find synergies with conventional approaches.
Abstract
Mistletoes have been considered a keystone resource for biodiversity, as well as a remarkable source of medicinal attributes that attract pharmacologists. Due to their hemiparasitic nature, mistletoes leach water and nutrients, including primary and secondary metabolites, through the vascular systems of their plant hosts, primarily trees. As a result of intense mistletoe infection, the hosts suffer various growth and physiological detriments, which often lead to tree mortality. Because of their easy dispersal and widespread tropism, mistletoes have become serious pests for commercial fruit and timber plantations. A variety of physical and chemical treatment methods, along with silvicultural practices, have shaped conventional mistletoe management. Others, however, have either failed to circumvent the growing range and tropism of these parasitic plants or present significant environmental and public health risks. A biocontrol approach that could sidestep these issues has never achieved full proof of concept in real-field applications. Our review discusses the downsides of conventional mistletoe control techniques and explores the possibilities of biotechnological approaches using biocontrol agents and transgenic technologies. It is possible that smart management options will pave the way for technologically advanced solutions to mitigate mistletoes that are yet to be exploited.
Keywords:
mistletoe; biocontrol; transcriptome; parasitic plant; resistance; seed dispersal; frugivore; environment; drone; remote sensing1. Introduction
Plants have been the principal source of food and energy for almost all living beings [1,2]. Economists and agriculturalists worldwide are concerned about food security due to growing populations and concomitant reductions in agriculturally cultivable land [3]. A variety of obstacles hamper efforts to improve food production, including human encroachment on land [3] and natural disturbances (forest fires, landslides, earthquakes, floods, etc.) [4,5]. In addition to the population-expansion-driven scarcity of agriculturally cultivable areas and vanishing forest ranges, biotic and abiotic stresses are equally serious and limiting factors for agricultural production [6,7]. Abiotic stresses include drought, temperature, cold, soil infertility, salinity, and the growing environmental concern regarding pollutants such as microplastics [8]. Biotic factors include overgrazing by higher animals, diseases from viruses, fungi, and bacteria, insect infestations, weeds, and parasitic plants. Most of these stressors, however, only episodically affect the growth of plants [9]. In contrast, parasitic plants such as mistletoes exhibit a prolonged presence, with many equating their senescence to that of their hosts [10,11,12,13,14,15,16].
There are nearly 5000 parasitic plant species belonging to 20 families of angiosperms, and many of them cause significant losses in crops used for food and industrial purposes [17,18]. Alongside the world’s major plant pests, Striga, Orobanche, Cuscuta, and mistletoes are more pronounced [16,19,20,21,22,23,24,25]. Striga, Orobanche, and Cuscuta have been reviewed extensively [19,20,21,22,23,24], but we would like to call attention to mistletoe, which has emerged as a growing global problem and requires better management and control [26].
The mistletoe is a hemiparasitic plant that clings to trees and depletes them of nutrients and materials, and, in many cases, heightened infection can result in tree death [27,28]. Efficient seed-dispersing mechanisms and/or frugivorous avians, as well as highly diversified haustorial structures, contribute to their enhanced tropism [27]. These pests severely affect tree plantations across the globe [27,29]. In addition to being a keystone resource of biodiversity and, to a somewhat minimal extent, their medicinal assets, they have also raised serious concerns regarding the commercial fruit and timber farming communities [28,30,31,32]. In spite of the efforts of these and some research communities, conventional mistletoe management approaches have not succeeded in mitigating the mistletoe problem. On the contrary, most of these approaches, such as deliberate fires, herbicide use, pruning, pollarding, plastic wrapping, etc., overlook damages caused to the environment and public health [27,29,33]. To counter this, newer approaches followed, exploring hyperparasitism from biological entities that thrive on the mistletoes and testing their worth as mistletoe biocontrol agents (MBCAs). However, to date, no MBCA formulation has yet been translated to the market. Despite some silvicultural trials in small-scale settings, other biotechnological interventions were limited to time-consuming and laborious tree-breeding strategies [34]. Beyond these, transgenic approaches and smart solutions have not yet been explored in the 21st century. In the face of emerging new host records and enhanced tropism, mistletoe seems to be outperforming management initiatives and losing the necessary pace of development and advancement.
This article examines the pros and cons of conventional mistletoe management and explores whether the biotechnological interventions have provided breathing room for biocontrol in terms of developing and using (i) biological mistletoe control agents and (ii) transgenic technologies to reduce the mistletoe burden on commercial fruit and timber tree plantations. The future possibilities with smart solutions for mistletoe management and the bottlenecks are also discussed.
2. The Biology of Mistletoe
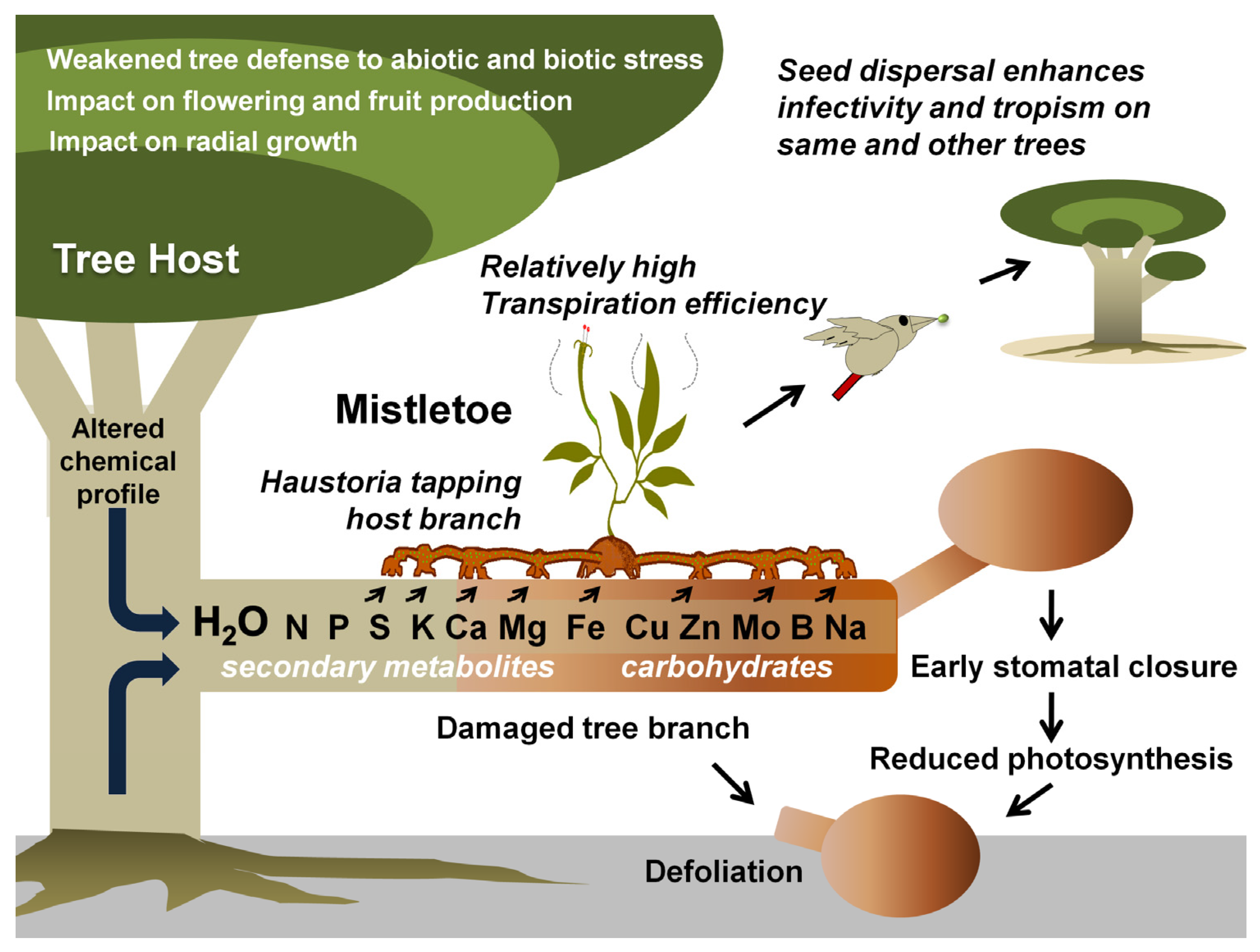
Mistletoes occur in the order Santalales, occupying the Loranthaceae (approx. 1000 species) and Viscaceae (approx. 550 species) [35]. Notably, the most notoriously damaging species in the Loranthaceae are the honey-suckled (Dendrophthoe spp.), the showy (Helixanthera spp. and Psittacanthus spp.), and the red mistletoes (Tapinanthus spp.), while, among the Viscaceae are the Dwarf (Arceuthobium spp.), the American (Phoradendron spp.), and the European mistletoes (Viscum spp.) [36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63]. Mistletoes are characterized as hemiparasitic plants because of their reduced photosynthetic efficiency and the absence of a true rooting feature [18,64]. A false root-like appendage, known as a haustorium, attaches them to their host plants (mostly trees) and draws water and nutrients from them [65,66,67] (Figure 1). Generally, these haustorial connections lack a retranslocation system, meaning that the hemiparasites directly and exclusively associate with the host xylem, but exploitation of the host phloem is never reported [68,69,70]. By transpiring almost nine times as quickly as their hosts, mistletoes suppress their host’s ability to maintain water potential, thus causing early stomatal closure and reduced carbon assimilation [11,64,71,72,73,74]. Hosts find it difficult to maintain their water, carbohydrate, and mineral profiles, especially under drought and soil infertility conditions [75,76,77,78]. In addition, climate change may add to the host detriments since mistletoe may spread to new geographical regions, possibly infecting new hosts and increasing in infectivity [71,75,76]. Mistletoe seeds are dispersed predominantly by fruit-eating birds [79,80]. Some birds have coevolved with mistletoes exhibiting fruiting displays [81]. Mistletoes attract a narrow range of avian dispersers that have anatomical adaptations and dietary preferences specific to mistletoe fruits [66,81,82]. Some Viscaceae mistletoe in the genera Arceuthobium and Korthalsella are equipped with explosive dispersal mechanisms in their seeds [66]. Seed dispersal by these modes has possibly allowed mistletoe to spread to nearby potential host trees, as well as those in islands and continents far off [83,84,85,86]. Seed dispersal on a compatible tree host marks the start of the mistletoe life cycle and parasitism (Figure 2).
3. Mistletoe Damages Trees
There is a great deal of liability associated with mistletoe in recreational settings, as well as commercial losses [26,28,31,90]. However, only a few surveys have been conducted at global scales to obtain data depicting the extent of damages caused by individual family members [87,91]. Several billion dollars’ worth of timber is lost each year due to dwarf mistletoe in Western Canada and the United States [92,93]. Many species causing heavy losses were reported early in the 1970s [94]. European mistletoe (Viscum album) itself is known to affect around 450 varieties of trees [95]. In North America and Canada, dwarf mistletoes (Arceuthobium spp.) predominantly affect the coniferous forests, especially tree species such as western hemlock (Tsuga heterophylla), lodgepole pine (Pinus contorta), and Douglas fir (Pseudotsuga menziesii) [51,96,97]. In Australia, eucalyptus and acacia plantations are heavily attacked by Amyema and Korthalsella species [98,99]. In Asia, the Korean mistletoe (Viscum coloratum), Macrosolen spp., and the Indian honey-suckled mistletoe (Dendrophthoe falcata) are more prevalent [100,101,102,103].
Depending on the parasite’s size, proliferation rate, and photosynthetic potential, as well as the host’s resourcefulness and developmental phase, mistletoe parasites have varying impacts on their hosts [44,46,59,60,61,104,105]. From the first infection, whether mistletoes kill the host or whether other factors offset this mortality is debated [28]. Many studies suggest that mistletoes negatively impact tree survival and fecundity, resulting in changes in population dynamics and structure, which in turn threaten their sustenance and conservation status [98,106,107,108,109]. An infected host (Figure 1) may indicate water stress and depletion of macro- (N, K, Ca, P, and S) [64,110,111,112] and micro-nutrients (Mg, Fe, Cu, Zn, Mo, B, and Na) [113], growth retardation [61,114], morphological and physical abnormalities [62,63,115,116], and shortening of stomatal opening [71]. Upon intensified infection, these effects slowly but eventually lead to the premature death of the host [39,60,117,118]. Besides the above direct damages, changes in the metabolic turnover and water stress (imposed by a competitively higher mistletoe transpiration rate, especially in water-limited scenarios) may render increased tree host susceptibility to microbial pathogens [106,119,120]. Infection with these hemiparasitic plants results in the loss of approximately 22–43% of carbohydrates in hosts [121]. Phoradendron perottetti affects Tapirira guanensis, resulting in the loss of many primary metabolites (especially soluble carbohydrates) and secondary metabolites (high levels of flavonoids) at the galls [53,61]. This is indicative of reactive oxygen species (ROS) quenching [53,61]. Further, a reduction in chlorophyll, foliage, and the biomass of hosts may reduce the photosynthetic potential as well as host biomass [52,58,59,62,122,123,124,125]. The radial growth of tree hosts has been reduced by Viscum spp. in Germany, Switzerland, and Spain [111,123,126], as evident from the reduction in annual growth rings in trees [125,127]. Moreover, mistletoe can interfere with flowering and fruiting and make the hosts more vulnerable to insect and fungal damage [128,129]. They also compete with host chemical profiles to remain healthy [130,131]. Lázaro-González and coworkers showed that Viscum album may load-dependently reduce the nitrogen concentrations in its pine hosts and may effectuate defense signals resembling drought, burning, and defoliation [130]. Recent work with Phoradendron crassifolium and Psittacanthus robustus revealed that tree branches neighboring the mistletoe infection site relatively experienced more adversities [132]. Studies have also reported a 40% reduction in mycorrhizal richness in tree hosts during mistletoe infestation [133,134]. In terms of foliar appearance, mistletoes often mimic their hosts [135,136,137]. This prevents them from being recognized by predators and herbivores that feed on them [135,136,137,138,139]. The exploitation of secondary metabolites from the host is known to confer such properties [135,136,137,140]. By doing so, mistletoe bypasses natural removal from hosts and escapes pruning or pollarding practices. The mimicry hypothesis in the case of Australian mistletoes, however, was questioned in a study by Blick and coworkers [141].
All the above detriments may result in the premature senescence of commercially valued tree varieties, such as fruits, timber, and others. Control of mistletoe is limited because of many contributing factors, including easy seed dispersal by frugivorous birds, explosive seed dispersal mechanisms, and reinfestation with remnants post-pruning, as well as others such as costs, labor, seasons, and climatic factors.
4. Conventional Control Strategies and Integrated Pest Management Approaches
Numerous conventional mistletoe control options have been routinely practiced by primary producers to mitigate mistletoe infestation. These include physical, chemical, and silvicultural approaches. We discuss the merits and downsides of these options below.
4.1. Physical Methods
Mistletoe was managed by people in forests and farms using various primitive methods. In the US, western dwarf mistletoe (Arceuthobium campylopodum) predominantly affects Ponderosa and Jeffery pines in forests [90,142]. These mistletoes can reduce life expectancy and result in the poor growth of infected hosts, as well as a loss of their scenic and esthetic value [90,142]. Conventional practices such as pruning and thinning of the affected host parts have proven less helpful [28,143,144,145]. There are many concerns and problematic issues that also have hindered these methods [90,145]. Known practices that have shaped the integrated management regimens for the control of mistletoes have been categorized into direct and indirect approaches [29,90]. Direct methods include the individual removal of infected trees, branches, or mistletoe brooms, or installing buffer strips, and the use of hosts that can resist mistletoe parasitism [90]. These methods could limit mistletoe spread and infection rates to some extent [90]. The potential benefit of removing the infected branches in Scot pines (Pinus sylvestris) aids in evading the inherent competition for water and nutrients within the host plant, which concomitantly results in improved tree health, such as elevated height, biomass growth, and diameter [111]. The downside of the direct methods, however, has been the labor and cost-intensiveness, limiting their application to only local small and high-value areas including parks, city limits, orchards, and nurseries [146,147]. It may be possible to remove large areas of mistletoe-infested trees; however, scheduling this around the growth periods of various hosts would be difficult and time-consuming [148,149]. Additionally, there are many modes of seed dispersal, including birds, winds, and storms, which make these strategies difficult. For example, it is difficult to expect a dwarf mistletoe-free stand within approximately a hundred feet of an infected Ponderosa pine [90,150,151,152]. Reid and Yan [153] highlighted the possibilities of developing aerial, ultra-light, surgical, and/or chemical delivery options. In either case, such approaches may not efficiently eliminate mistletoes from heavily parasitized trees [153]. Surgical methods require time, labor, and financial costs, making them impracticable for small- to large-scale plantations [29]. Owing to expenses, farmers inevitably cannot employ professional services from arborists and tree surgeons who offer tree lopping and pruning options [29]. For them, the only option is to resort to the age-old traditions of pollarding, as in Southeast Australia [29]. This, in addition to mistletoe control, gives them fodder during droughts, shade for livestock, firewood, and fencing [29]. Other physical methods, such as the use of paints and deliberate fires, are viable measures [154]. Fire applications have offered some appreciable control of the mistletoe population in the western USA forests [155,156,157], because of mistletoes’ slow recovery rate compared to host(s) [158,159,160]. However, the use of fire may have limitations in that a weakened host may favor mistletoe regrowth [51,161,162]. Moreover, on holistic grounds, fire applications may raise public and environmental health concerns [163,164]. Indirect methods use irrigation, fertilization, brush and weed control, and regulating human disturbances [29,90,165]. Irrigation profiles of the host may variously impact hemiparasite infectivity. In many cases, the higher water status of the host corroborates with heightened mistletoe infectivity, hinting that intermittent drought regimes would offer some protection for the hosts [90,166,167,168]. Improving the fertility of hosts by the use of nitrogen fertilizers has shown some protection from the mistletoes’ effects, with increments in tree height [169]. Other indirect approaches also aid in improving host vigor and thus can be applied in large settings [90]. Above all, scientific evaluation of the above strategies requires surplus funds and support for research and development communities to devise containment facilities and flexibilities in regulations to work in protected forest areas [29].
4.2. Chemical Methods
Though with only limited prospects, chemical-agent-based approaches to mistletoe control have been more prevalent in various non-European countries such as India, Bangladesh [57,170], Australia [171], and the United States [88]. These methods involve the use of sprays or trunk injections of certain herbicide formulations to selectively control the hemiparasite without negatively impacting the host plant species [171,172,173,174]. In Eastern Australian farm eucalypts, the use of 2,4-D formulations had been investigated long ago in the 1950s and 1960s to control box mistletoes (Ameyema spp.) [174]. Besides their poor control, the trials could not be widely extended or reproduced with equivalent success due to the difficulties in ascertaining the optimum dose as influenced by various factors such as mistletoe host species, its size, the season of injection, the frequency of mistletoe infestation, and others [171,172,173]. Formulations such as 2,4-D, 2,4,5-T, 2,4-MCPB, and dichloroethane have substantially been utilized to destroy Viscum album subsp. abietis on Abies with only insignificant host effects [34]. However, complete resistance to mistletoe is never induced in hosts, as seen in the case of using 2,4-D and glyphosate, wherein mistletoe control only lasts half a year post-treatment [34,175]. A more promising herbicide control measure, according to a large study by Quick [176], involves using an iso-octyl ester of 2,4,5-T, which was later banned for its non-target effects [34]. It was shown that numerous other formulations tested between 1970 and the mid-1990s, such as Nutyrac, Dacamine, Thistrol, MCPA, Goal, D-40, Emulsamine, DPX, Prime, and Weedone, were unreactive to the endophytic system, despite killing mistletoe foliage exclusively, without adverse effects on the host [128]. Similarly, ethephon (Florel, 2-Chloroethyl phosphoric acid), which releases ethylene, a plant growth regulator, has been tested with various dwarf mistletoe–host models in Manitoba, California, Idaho, New Mexico, Oregon, and Minnesota [177,178,179]. With few non-target effects, it results in defoliation in mistletoe, albeit without influencing the haustorial connection, meaning that the complete evasion of infection is still a concern and recurrence cannot be ruled out [180]. A defoliated but active haustorium can persist for over a century [87].
It is still necessary to ensure the prompt delivery and adequate coverage of chemical agents, even if they appear potent enough [26,34,181]. Ground applications can be effective but may not reach the hemiparasites in the tree overstory higher up in the canopies [34]. Helicopter-based aerial delivery approaches, however, are challenged by poor penetration to the lower crowns, as sprays would be hindered by the overstory crowns [182,183]. To continue, chemical control methods pose many difficulties, such as limited technologies for the selective treatment of mistletoe over hosts, chemical damage to hosts, unavoidable reinfestation due to parasite seed germination or spread after chemical application, developing crops with a high tolerance to chemical applications, low penetration, persistence, availability, and so on [180,184]. Again, chemical agents that do not harm the haustorial connections may gradually exhibit resprouting [185]. This will be graver in areas with greater incidences of mistletoe infection and its resprouting-led intensification [185]. Despite the best chemical agents, mistletoe seeds will only be delayed for 2–4 years, so there is only a temporary solution to the problem [34]. Considering these factors, alternative control and management methods are highly sought after.
4.3. Silvicultural Practices
In contrast to the above approaches, silvicultural practices have been more promising [90,128,148]. These work by first clearing mistletoes from infection sites and then reintroducing a fresh lot of plants to the area [186]. Such methods have been implemented for controlling dwarf mistletoes in North America [128,186]. Robinson and colleagues proposed a spatial–statistical model that simulates mistletoe growth and development under many control methods and offers a more preferable option [187].
In small recreational and commercial settings, a silvicultural approach focuses on multiple factors, including ecological disturbances, harvests, prescribed fires, and need analysis and impact assessments [148,186]. They have gained acceptance as a routine strategy for highly devastating mistletoe and have slowed the development of resistant cultivars [34]. Silvicultural practices, however, may not guarantee long-term and consistent efficacies due to the gradual changes in climate and shifts in devastating mistletoes related to their enhanced tropism [186]. In order to scale this approach and ensure its success, newer options are highly desirable. A recent investigation showed an associational resistance to the spread of Viscum album ssp. austriacum in Northern Spain and indicated that mixed plantations of hosts (Scot pine, Pinus sylvestris) with non-host (Maritime pine, P. pinaster) trees greatly reduce mistletoe infestation [188].
5. Mistletoe Control through Biotechnological Interventions and Smart Management
Biotechnological approaches and the use of 21st-century technological advancements can bring a paradigm shift to the current mistletoe management regimens. Mistletoe control biotechnology should realize the use of potential MBCAs, which variously deteriorate and kill the mistletoes on tree hosts (Supplementary Table S1). Alternatively, enhancing or introducing traits conferring mistletoe resistance in host trees should be considered a more promising option [189]. The MBCA approach has seen limited application in finding and characterizing potential microbial and insect pathogens, as well as other life forms, but is yet to see a fully mechanized industry setup and global application [190]. The genetic approach similarly relies on mere conventional breeding trials [128,191]. In the sections ahead, we examine the current status of these highly promising biocontrol options and develop various plans to take advantage of them.
5.1. Mistletoe Biocontrol Agents (MBCAs)
Organisms with specific nutritional requirements that jeopardize their concomitant host(s), have emerged as a biocontrol strategy in most pest management routines [24,192,193]. Integrated pest management practices have become an established part of agriculture, forestry, and horticulture [194]. Such biocontrol agents offer wide acceptability and ease of application due to their straightforward and easy production processes and various biosafety and eco-friendly attributes [195]. In particular, for use as MBCAs against dwarf mistletoes, various fungi, bacteria, and insects have been proposed [196,197,198,199,200].
5.1.1. Bacteria and Fungi as MBCAs
Many well-known marketed mycoherbicidal formulations are available as MBCAs for controlling woody weeds, viz., DeVine® (Phytophthora palmivora for controlling strangler vine on citrus) [201], Collego® (Colletorichum gloeosporioides for controlling Northern Jointvetch in rice and soybean crops) [202], and BioMal® (Colletotrichum gloeosporioides for controlling round leafed mallow weed in agricultural settings) [203]. Chondrostereum purpureum, a primary wood invader, has been investigated by Canadian and European researchers as a biocontrol solution for undesirable woody weeds in conifer regeneration and forestry settings [204,205,206,207]. Similarly, StumpOut®, a registered biocontrol product, is a white-rot fungus (Cylindrobasidium laeve) used in South Africa for controlling Australian wattles [208]. As with weeds, hemiparasitic mistletoes also harbor a range of microbial pathogens with varying species specificity. Hyperparasitism from both fungi and bacteria (hence myco- and bacterio-herbicides, respectively) can be used for mistletoe biocontrol, some of which are listed in Table S1 (Supplementary Material). Fungal pathogens reported on many Australian Loranthaceae mistletoes [209] receive proper attention regarding their potential as MBCAs. To explore control over dwarf mistletoes, the myco-MBCA candidates are considered in two categories: the aerial shoot fungi that act on the foliage, and the canker fungi, which attack the endophytic system [210]. More than 20 different canker types are known for Arceuthobium tsugense in British Columbia; however, because their MBCA potential includes both pros and cons, they require in-depth assessments before mass field application [87]. Nevertheless, three potent canker fungi are described. American and Canadian dwarf mistletoes (Arceuthobium spp.) commonly harbor a fungus, Colletotrichum gloeosporioides, which has been investigated for its potential as a mycoherbicide MBCA for controlling this mistletoe on western hemlock and lodgepole pine [210]. This fungus is virulent, easily cultured in vitro, and completely disrupts mistletoe growth shortly after shoot emergence, and it has shown success in field trials attempted in British Columbia. Especially for A. americanum and A. tsugense, in vitro assays have been developed to evaluate the potential of this fungus as an MBCA [211]. Other than this, a canker fungus, Neonectria neomacrospora, has also been isolated with documented pathogenicity from cankers caused by A. tsugense in British Columbia [212]. In addition to its ability to infect its mistletoe host without wounding, quick sporulation, selective spread, foliage-reducing properties, and mortality characteristics, the fungus is selective to infect dwarf-mistletoe-infected host tissues. At field sites for dwarf mistletoe, intensive efforts are being made to optimize and improve industrial-scale production and delivery methods for this MBCA [213]. The anamorph-staged fungi Cylindrocarpon has been tested in vitro as another, yet more promising MBCA (than C. gloeosporioides), with aggressive colonization on dwarf mistletoe seedlings and callus [211]. Cytospora abietis Sacc. is a well-known fungus attacking dwarf mistletoe cankers on red and white fir species Abies magnifica and A. concolor, respectively [214,215].
Arceuthobium americanum, parasitizing Pinus contorta of the Rocky Mountains in Northwestern America, commonly succumb to Resin Disease Syndrome, a disease outcome of a complex attack from various fungi, which predominantly includes Alternaria alternata, followed by Aureobasidium pullulans and Epicoccum nigrum [216]. This can result in excessive resinosis of the haustoria, necrotic lesions in the bark, and dying foliage as symptoms potentiating MBCA candidature for each of the pathogens. However, these fungi have also been isolated individually from non-symptomatic mistletoe–host associations, indicating that the syndrome is a multipathogenic phenomenon [48]. Substantiating an MBCA, potentially of individual fungi or one MBCA cocktail formulation, hitherto demands further investigations on reproductive potential, the comparison of systematic and non-systematic infection in mistletoe, and the characterization of environmental and fungal components as well [216].
Viscum album negatively affects a variety of pine trees in Turkish forests, which are important timber crop species [105]. Kotan and coworkers [217] analyzed the MBCA efficacies of several bacterial and fungal isolates from the foliage of various diseased V. album. They reported five possible bacterial MBCAs [217], which were pathogenic only when administered to mistletoe through injection, and not when sprayed. In addition, they reported four highly pathogenic fungal strains that showed potential for MBCA when sprayed. Very recently, Aureobasidium harposporum, a new fungus, has been isolated, which causes leaf spot disease on V. album in Turkey [218].
5.1.2. Diatoms and Algae as MBCAs
According to reports, diatoms and marine algae resist parasitic bacteria, fungi, and insects [219,220,221,222,223,224]. However, very little is known about their efficacy in controlling parasitic plants. In a recent study, a commercial diatomaceous earth formulation, Mistletoe Killer®, was reported to parallel ethephon in controlling the mistletoe Arceuthobium globosum ssp. grandicaule parasitizing Pinus pseudostrobus [47]. Accordingly, a 7.5% dose of this formulation resulted in foliar death and complete mistletoe removal (respectively, after 15 and 45 days of application), without impacting the tree host. However, follow-up studies aimed at understanding the basis of the selective damage to mistletoe from diatoms remain to be pursued. It also remains to further test this formulation on other mistletoe–host pairs, which could validate the MBCA potential of diatoms and other algae’s application.
5.1.3. Insects as MBCAs
Several insect classes have the potential to become successful MBCAs. In 1984, J.D. Solomon, an entomologist, reported a stem borer weevil, Myrmex sp. (Coleoptera: Curculionidae), from the declining clusters of the mistletoe Phoradendron serotinum occurring on many water oak trees (Quercus nigra), near Stoneville, Mississippi [225]. This weevil tunnels into the mistletoe stem and deposits its larvae, which pupate and emerge, making holes and galleries within the stem, leading to mistletoe diebacks upon heavy weevil attacks. Before this, other species in this insect genera, M. arizonicus, and M. algerti, were also reported from mistletoes in the United States [226,227]. Recently, another phytophagous weevil, Timorus sarcophagoides (Coleoptera: Curculionidae) [228], has been reported to feed and carry out its life cycle exclusively on the parrot flower mistletoe, Psittacanthus robustus (Loranthaceae). It feeds on soft tissues in the flower buds, pollen grains, stamens, ovaries, and leaf axils, as well as digging tiny holes in the haustoria to oviposit and lay eggs. Adults are known to walk throughout the mistletoe and rarely leave it. Moreover, by closely resembling flesh flies (Diptera: Sarcophagidae), these adults represent a valuable example of evasive mimicry, enabling them to be overlooked as uncatchable prey by birds and other predators [229]. For use in MBCA, this property may ensure extended consistency of application.
Loranthus spp. predominantly attack tea, citrus, guava, rubber, kapok, rubber, and eucalyptus plantations in many parts of the world [87]. In two old monographs documenting a cursory survey of phytophagous flora in West Pakistan, Mushtaq and Baloch have reported many species of insects and mites infesting Loranthus longiflorus [230,231]. These researchers also explored the mistletoe specificity in insects to account for their suitability as an MBCA. Of these insects, some were never reported for any alternate host, while others were more destructive to the tested mistletoes, proving their worth as putative MBCAs. These were oligophagous in their restricted inhabitance to Loranthaceae mistletoes.
Some insects complete part of their life cycle on mistletoes in the form of egg deposits [225,226,227,228]. One such family is Buprestidae (Coleoptera) of the Jewel beetles [232,233]. They include Agrilus viscivorus, A. graceus, A. jacetanus, and A. kutahyanus [232,233], all of which thrive on mistletoe sap and show a detrimental influence over the mistletoes, eventually killing them upon their maturation.
Moths, such as Synanthedon loranthi (Lepidoptera: Sesiidae), have also been found harboring mistletoes that infect Scots pine [234]. The larva burrows into the mistletoe stem and pupates on the leaves [234]. Some moths belonging to the family Tortricidae, such as Celypha woodiana and Ditula angustiorana, feed over the mistletoe leaves [235,236,237]. Hemipteran moths such as Cacopsylla viscid, Anthocoris viscid, and Pinalitus viscicola have also been reported on mistletoes [238,239].
In another case, scale insects have been evidenced to cause disease symptoms over mistletoes. False oleander scale insect (Pseudaulacaspis cockerelli; Hemiptera: Diaspididae) thrives over the honey-suckled mistletoe (Dendrophthoe falcata var. falcata; Loranthaceae) found parasitizing the leguminous Cassia trees (Senna siamea) in India [37]. Other than this, a cottony cushiony scale insect (Icerya purchase; Hemiptera: Monophlebidae) was reported exclusively to be infective on the red-berried mistletoe (Viscum cruciatum), and not the mistletoe host, the olive tree [240]. Importantly, tree hosts in both cases were uninfluenced by scale insects, meaning that the latter could be selective to mistletoes.
There is a need to carry out faunistic surveys and sampling on various mistletoe–arthropod associations so that enough resources are available for lab-to-field-scale tests before reaching a conclusive remark on the candidate MBCAs of the insect world [241]. Similarly, many of the previously reported insect fauna from the New World dwarf mistletoes in Asia require detailed studies [242].
5.1.4. Hyperparasitic Mistletoes
Some mistletoe species may parasitize other mistletoe species growing over tree hosts. This is termed hyperparasitism [243]. For example, the Bollean mistletoe (Phoradendron bolleanum) usually grows on trees (in the genera Juniperus and Arbutus), but occasionally appears as a facultative hyperparasite on various mistletoe species [244]. Viscum capitellatum in Sri Lanka is an obligate hyperparasite of widespread mistletoe Dendrophthoe falcata L.f. [245]. There are many examples of hyperparasitism in both Loranthaceae and Viscaceae [55,246,247,248]. Plant phenology and seed dispersal agents’ behaviors play important roles in the chance occurrence of hyperparasitism [243]. However, developing such MBCAs would be illogical and difficult in managing (i) the technical diligence of their delivery for controlled release and (ii) off-target effects and concomitant outbreaks. In addition, it may switch to parasitizing the host tree, which would otherwise be rescued and restored from mistletoe attacks. The realization of such an approach would also require a deeper understanding of mistletoe’s compatibility with hosts, epigenetic influences, and other factors.
5.1.5. Higher Animals
Higher-order organisms in the animal kingdom may aid in devising mistletoe control measures [29]. Common bushtail possum (Trichosurus vulpecula) is a leaf-loving marsupial native to Australia [249]. In a conference proceeding by Sessions and co-workers, this mammalian species was reported accountable for the decline of the endemic Loranthaceae mistletoes Peraxilla tetrapetala, P. colensoi, and Alepis flavida [250]. Around 50% of the studied Peraxilla mistletoes were defoliated by the possums between 1978 and 1982 at Australia’s Nelson Lakes National Park [250]. This could pave the way to a newer strategy to use such animal species in countries where mistletoes heavily damage commercial fruit farms and forests [251,252]. However, this control option could be challenged by the declining habitats for these animals onsite [25].
5.2. Requirements of an Effective MBCA Selection Program
A comprehensive investigation of MBCAs may require expansive sampling and surveys towards generating metadata on biological entities, viz., insects and microbial communities that selectively impose pathogenic symptoms on mistletoes without harming their respective tree hosts. Future research should focus on individual entities and on characterizing the performance of emerging MBCAs against specific mistletoes. There is a need for rigorous research on the synergistic effects of two or more potential species (which could create an MBCA cocktail) for more notorious mistletoe(s), areas complexed with more than one variety, or for those that attack multiple hosts at the same time. Examples of primitive studies of this type exist [253,254]. Insects have equal potential as microbes for use as MBCAs, as many in their larval form feed exclusively on mistletoes [230]. In terms of MBCA potential, mistletoe pathogenicity alone cannot provide a concrete solution. Before selecting MBCAs, a checklist of features that best suit the testing of putative entities should be developed. An ideal checklist should include all the possible critical attributes on which MBCA hyperparasitism can be tested on acid, with a scoring system for their efficacy inherently (as MBCA) and their impact on the mistletoe, its tree host, and other non-host plantations. MBCAs would also require mass production, effective delivery systems, deployment strategies [34], and a consistent monitoring program. Below is a comprehensive checklist of parameters that can be used to test and screen MBCA candidates (Figure 3).
5.3. Challenges with MBCAs
A different perspective considers pathogenic microbes and insects as co-evolving complex relationships with mistletoe hosts [34]. Mistletoe outbreaks are induced or regulated by a variety of factors associated with seasonal and weather variations and their influences on multitrophic communities [34]. This emphasizes the importance of intensive surveying, isolation, and study of the mistletoe–host specificity for each of the biological entities carrying considerable potential as an MBCA. Perhaps many microbes may appear circumstantially sensitive even to minute changes in their growth parameters [256,257]. The culturability and preservation (and conservation) of such organisms is also very important for reproducibility in continued/extended studies and applications. Many of these might be new, yet unexplored, rare, or others nearing extinction regarding strains, varieties, species, and of course their MBCA attributes. Moreover, some of these putative MBCAs may be lost due to environmental calamities and/or seasonal variations, especially for those very specific to the endemic mistletoes (especially their varieties). This must hold for entities that have no alternate host(s) than mistletoes. Examples of this could be the fruitfly genus, Ceratitella, represented by only four species, all of which thrive exclusively on mistletoes [258], or moths such as Zelleria loranthivora (Supplementary Table S1). Potential MBCAs are rare due to the scarcity of hosts that would otherwise enable their ecological expansion on more than one or very few mistletoe species. This seems imperative, especially for those endemic to not-so-vast geographic areas, where MBCA entities might lose some of their importance and worthiness if physical and chemical treatments are practiced in these areas. This will be more serious for entities that are closely related to, dependent on, or paralleled with mistletoe hosts. Mushtaque and Baloch’s study, for example, could not test six of the twelve insect species for MBCA potential [231]. The conservation and production of rare isolates may represent new technical challenges, such as strain- or species-specific media compositions, microbial culturing, and insect-rearing protocols. In addition, selecting MBCAs would pose the challenge of competing with other members of the host–mistletoe microbiome, as has been highlighted recently [259].
5.4. Inducing Host Plant Resistance to Mistletoes and/or Herbicides
Inherent resistance to mistletoes in some trees, such as Chinese pistachio (Pistacia chinensis), crape myrtle (Lagerstroemia), sycamore (Platanus occidentalis), and conifers such as cedars (Cedrus) and redwood (Sequoioideae), has been documented amongst their susceptible genotypes [260]. A call to develop a controlled breeding program catering to the identification of genetic bases for inherent resistance in tree hosts to hemiparasitic mistletoes was made as early as the 1960s [189]. Due to the advent of cost-effective and convenient silviculture solutions, it has received little attention from the scientific community [128]. Understanding the interactions of host–mistletoe compatibility would allow the selection of resistant varieties. Conventional breeding strategies, nevertheless, have developed some stably resistant tree lines [261]. Tree hosts may be evaluated for resistance to herbicides, which may aid in the better application of chemical control approaches following herbicide translocation through such trees to their mistletoe parasite(s). For many tree crops, breeding programs are time-consuming, labor-intensive, require extensive field trials, and have not resulted in herbicide-resistant cultivars [262]. Moreover, the chemical treatment of mistletoes on herbicide-resistant tree hosts might lead to the emergence of herbicide-tolerant mistletoes.
5.5. Hunting for the Genetic Basis to Hosts’ Inherent Resistance to Mistletoes: Background Studies
Because mistletoes have been co-evolving with their tree hosts for nearly 25 million years [263], the emergence of inherent resistance in these hosts cannot be denied [264], especially for the very native and devastating mistletoes [265,266]. A genetically directed effect is also implied by the fact that mistletoes exhibit specificity to their concomitant hosts, as well as variability in host preferences [36,64,165,247,267,268,269,270,271,272,273,274,275,276]. For example, Arceuthobium douglasii does not parasitize Pinus ponderosa [128]; around 70% of dwarf mistletoe species with a principal host also pathogenize other hosts and still with variable levels of symptoms [191]; and A. pusillum fluctuates in terms of infection extent when exposed to Larix laricina, Picea glauca, P. rubens, and Pinus strobes [277]. On the contrary, there are also reported instances where mistletoes, in a heavily infected area, skipped infecting their principal host. A few observations were reviewed previously for Arceuthobium spp. [34] and one for Dendrophthoe falcata more recently [36]. However, studies are yet to present convincing demarcating features of preferred and not-so-preferred host type(s). These reports suggest existing variations in exhibiting resistance within the host population, even though the progeny of these trees have not been tested for resistance (a hotspot for further investigation). It is evident from these findings that few data are available regarding species-specific susceptibilities, which calls for rigorous field examinations such as progeny testing. Grafting techniques with putative resistant tree hosts used the inoculation of grafts and/or out-planting in mistletoe-infected sites [265,278]. Based on these trials, it appears that resistance is genetic rather than environmental. However, the heritability of this genetic regulation could not be documented. Many progeny tests to elucidate the heritability initially met with mixed results, with some escape or non-inheritable events [128,279,280,281]. Positives incidences [265,266,282,283] and later isozyme data from Nowicki’s work (as reviewed in Shamoun and DeWald [34]) more strongly support heritability.
5.6. Mistletoe Community Restructuring, Disturbances, and Biological Interactions
Mistletoe infestations drastically differ at various geographic locations, and as such from urban to forest settings [275,284,285,286,287]. Their prevalence in these areas can be variously influenced by the occurrence of host species [276], host specificity [275,285], the behavior of dispersing agents [79], pollinators [288], habitat disturbances [154,289], herbivory [138], and chemical interactions with the hosts [290,291]. Mistletoes may be differentially prevalent and infective from one to another host and within a population(s) of hosts [292,293]. Generalist mistletoes have a broad host range, while specialists exhibit host preferences [64,109,275,294]. Moreover, community dynamics profoundly impact parasite–host relationships [79,80,108,269,270,295]. Numerous life forms interact directly or indirectly with mistletoe, exhibiting either generalist or specialist behavior in doing so [40,79,80,81,83,84,296,297,298]. Thus, field surveyors must consider influences under various circumstances before taking into account mistletoes’ infectivity, host preferences, and inherent resistance in the hosts within a particular geographical area (especially dense forest settings and commercial tree plantations). Vegetation disturbances such as the incidences of dead trees, grazing, logging, forest fires, and human waste disposal, among many others, may both positively and negatively affect mistletoe prevalence [50,299,300,301,302]. Other than these, mistletoes may differ in their ability to readily establish seedling growth and mature persistently on various hosts [273,303,304]. Mistletoes such as Psittacathus calyculatus, Dendrophthoe falcata, and Phoradendron californicum associate readily with leguminous tree hosts [37,39,305]. Mistletoes’ diversity and evolution may be effectuated by host shifts, followed by host specialization, as well as variations in climate and environment [306]. In addition to these factors, mistletoes’ success as parasites also depends on their complex interactions with herbivores, pollinators, and seed dispersers, such as birds, insects, and bats [40,41,298,307,308,309,310]. Many also believe that the genetic structuring and isolation of mistletoe into populations growing on various hosts within a community is attributed to these avian gene dispersal vectors [41,42,311]. However, based on the notion that most of these agents are insufficiently specialized, some believe that mistletoe speciation is mainly driven by host–parasite interplay [274,296,297,306,312,313,314]. Inconsistencies in mistletoe frequency within host populations are primarily due to seed dispersal mechanisms and disperser behavior. Some mistletoes (such as Arceuthobium spp.) exhibit explosive or ballistic seed dispersal [83]. Bird-dispersed mistletoes generally show a patchy distribution, which is attributable to birds’ eating, roosting, and nesting behaviors [39,54,315,316,317]. Forests with endemic marsupial seed dispersers may hold a similar analogy for effects on mistletoe prevalence and distributions [296,318,319,320]. Many host factors, such as tree diversity [86,273], height, crown width [43,49,295,321,322,323], and bark type [267], as well as the recently studied stand characteristics [105], regulate mistletoes’ colonization and density. These factors may also variously influence seed disperser behaviors. For example, the Sonoran desert mistletoe (Phoradendron californicum) seeds are reported to be less frequently deposited on host tree species Cercidium microphyllum and Acacia constricta [305]. The dense, thorny, and entangling crowns in these trees probably do not facilitate the perching behavior of avian dispersers [305]. Additionally, competition among the host trees may reduce resource availability and negatively impact mistletoe infection and occurrence [78]. Hosts may influence the reproductive phenology of their mistletoe parasites [324]. This could be indirectly orchestrated by host-mediated influences over the pollinator communities with distinct pollinator rewards and those over mistletoes with pollen receipts, as well as avian visitations from distinct taxa on distinct hosts [325,326].
Besides influencing the interactions above the ground, mistletoes also tend to restructure the communities in the soils underneath their hosts and act as facilitators of heterogeneity, especially in low-productivity areas [327,328]. They may contribute to a more nutrient-rich litter from the abscission of their leaves, fruits, flowers, and seeds, which decompose faster and enhance soil-nutrient cycling. The litter may pave the way for newer forms of community interaction. For example, Hódar’s group reported that Viscum album litter may favor herbaceous vegetation and frequent visitations by rabbits underneath their host trees [329]. Another study shows that mistletoe-remodeled soil litter can improve habitat quality for insectivorous birds that prefer specific arthropod populations underneath mistletoe host trees [330]. As a result, nearby tree hosts (mistletoe-preferred and non-preferred) become more competitive. Other than these, mistletoes can generate non-tropic and trait-mediated detrimental indirect interactions with insects sharing the same tree host [331]. The mistletoe can therefore affect the entire herbivore community and their dependents, such as insectivorous birds and other plants, some of which may serve as seed dispersers and pollinators. Alternatively, mistletoe infection may, directly and indirectly, influence defense systems in their hosts to natural enemies. This has been reported in some pine trees exhibiting resin duct defense against bark beetles [332]. Mistletoe-inflicted changes to host chemical profiles may even influence insect behaviors and drive their evolutionary transition to other hosts. This is exemplified in a study that reports the role of dwarf mistletoe (Arceuthobium spp.) in mediating butterflies’ (Nephasia spp.) interaction with their pine hosts [272]. Climate change, on the other hand, also may variously impact the global distributional shifts in mistletoe [166,167,333,334]. These factors likely impede field investigations that could otherwise identify genetic controls of mistletoe resistance. In this case, specially designed containment facilities such as glass or net houses may offer robustness to field studies. Moreover, research on heritable genetic factors is lacking in the literature. It is still unclear whether the resistance phenotype in the host is governed by specific genes and/or is a cumulative effect of genes encoding other traits. Some studies hint at the latter possibility of resistance resulting from anatomical changes in the response to aging and wounding [265,277,283]. Alternatively, exploring the possibility of genetically evolved secondary metabolites in resistant hosts for defense against mistletoes also might be promising [268]. These represent gaps in existing studies, especially regarding the functional genomics of mistletoe–tree host associations, which could translate into their genetic manipulation and engineering for stably resistant transgenic trees. We discuss these opportunities and others in the sections ahead.
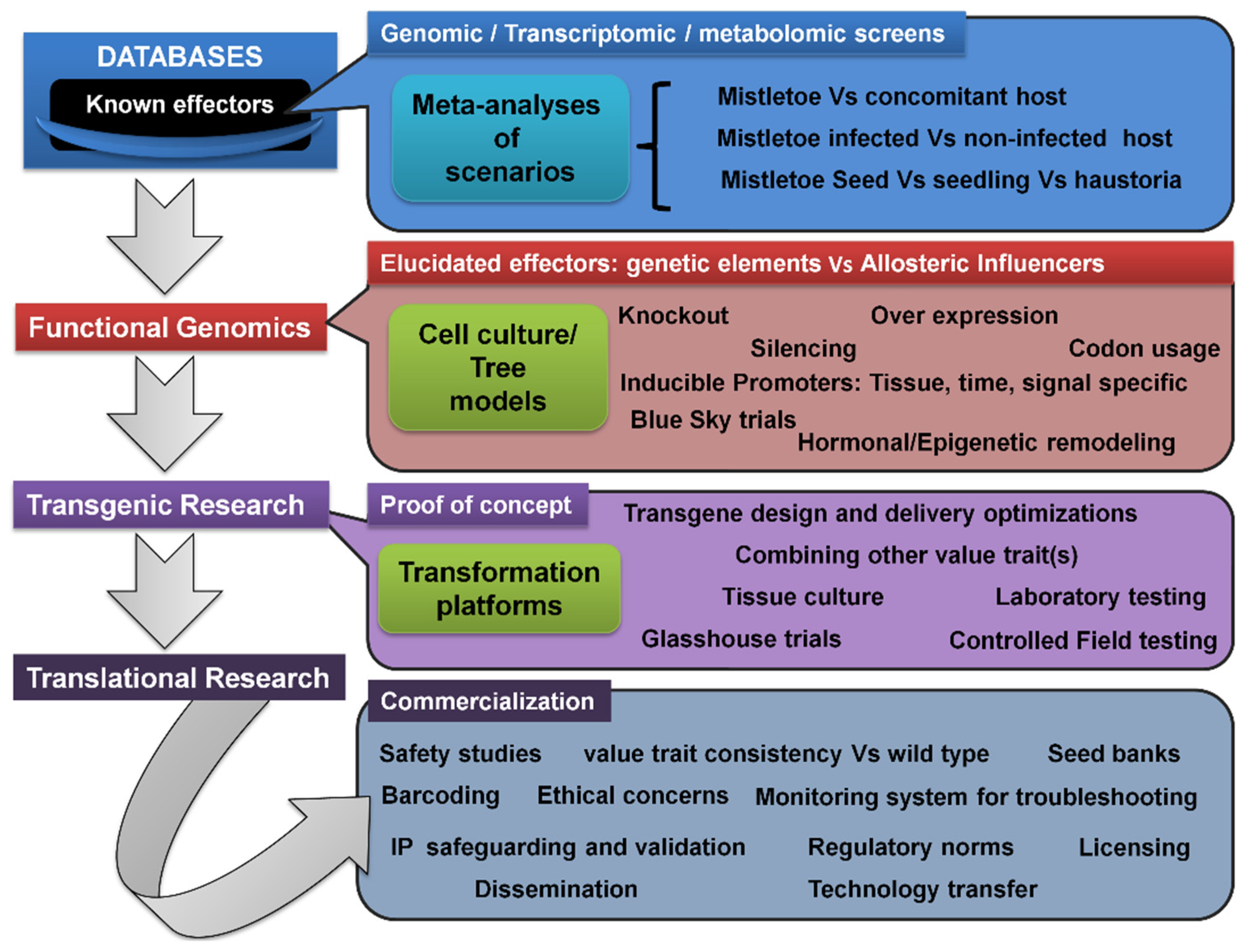
5.7. Transcriptomic/Metabolomic Profiling, Transgenic Trees, and Translational Research Pipeline
The genetic modification of mistletoe hosts to induce resistance, resulting in a new variety, could be a promising strategy for reducing damage [335]. This would require the development of screening programs to characterize the viable sources of host resistance to each of the mistletoes (Figure 4). Such screens can be examined for molecular DNA tags, aiding in expedited marker-assisted selection for mistletoe-resistant hosts. Nonetheless, limited but considerable progress has been made in this direction [34,336,337,338,339]. Metabolomic profiling in other parasitic plants indicated the upregulation of auxin-responsive genes at the host–parasite interface [340,341]. Ichihashi’s group recently carried out both transcriptomic and metabolomic profiling of haustoria in the root hemiparasite Thesium chinense, revealing upregulated transcripts for genes involved in the biosynthesis and signaling of very long-chain fatty acids, auxin, and lateral rhizogenesis [342]. Along similar lines, Wang’s group attempted tissue-specific transcriptomics and whole-genome expression profiling of the shoot, flower, fruit, and seeds of spruce dwarf mistletoe (Arceuthobium sichuanse), revealing 22,641 differentially expressed genes related majorly to photosynthesis, carbohydrate and cell metabolism, transcription, hormone biosynthesis, and their signaling [343]. Recently, Wahid’s work extensively used tools such as suppression subtractive hybridization (SSH) and next-generation sequencing (NGS) to report some 1279 genes in Ziarat junipers (Juniperus excelsa) in Balochistan, which might putatively confer resistance to the dwarf mistletoe Arceuthobium oxycedri [344,345]. Transcriptome profiling of Taxillus nigrans individuals on four different tree hosts revealed many host-specific and common pathways driving this mistletoe’s parasitism [346].
Research into transgenics would require a greater understanding of mistletoe’s genetics and molecular basis, as well as the dynamics and kinetics of materials’ flow between the host and its parasite (Figure 4). Alternatively, where the same mistletoe parasitizes multiple hosts, it might be interesting to identify similar genetic and/or molecular elements, their influences, and their interactions within these frameworks. In addition, a comprehensive comparison of meta-analyses arising from infection instances between the same host and different parasite pairs could provide insight into rigidities, flexibilities, crucial events, genetic elements, and/or their encoded influences within these frameworks. These characterizations may lead to the development and testing of transgenic host(s) that are resistant to mistletoe attacks, but also produce fruit and timber in a qualitative and quantitative manner. Technically speaking, genetic engineering is endowed with certain choices of gene knockout and gene silencing options. In addition to site-directed mutagenesis, RNA antisense, small interfering RNA (si-RNA), and the CRISPR-Cas9 approaches, there are time-tested ways to manipulate transcription factors [347,348,349,350,351]. The use of transgenic tree crops for translational research (Figure 5) would revolutionize and open new avenues for commercial fruit and timber businesses worldwide, eliminating the need for the cost-, time-, and labor-intensive management of mistletoe pests, and potentially ending primitive conventional methods that are not eco-friendly. Other than this, transgenic approaches may be combined with chemical control options. This may be realized by developing hosts that can withstand the otherwise mistletoe-intolerable levels of herbicides while delivering them to deter mistletoe hemiparasites. In this arena, we believe that much of the background work has already been done, as in Oronache and Striga [262,352], and can also be seen in the launch of a transcriptome meta-analysis initiative, the Parasitic Plant Genome Project [353]. There are a variety of reports that offer foundational studies on tree functional genomics [354,355,356,357,358,359]. As highlighted before, biotechnological approaches such as genetic engineering and plant tissue culture [211,360] have been underutilized, which indeed could represent a step away from conventional breeding strategies. There are few examples where tree transformation has been attempted in this regard. This should be seen as a critical requirement for introducing the usefully valued trait into specific fruit and timber tree cultivars [361]. In mitigating the mistletoes, however, such initiatives are yet to see fruitful realization and technology translations.
5.8. Role of Epigenetic Signaling in Mistletoe Parasitism: Moving beyond the Genetic Basis
It is also important for scientists to determine how mistletoe parasitism is epigenetically modulated or influenced. Mistletoes exhibit epigenetic influences in their foliar appearance, mimicking their hosts [362]. We have previously reported the unusual behavior of the loranth Dendrophthoe falcata var. falcata, exhibiting two different haustorial types on different fruit tree hosts [36]. Moreover, in areas where more probable and notable hosts were present, only a few uncommon hosts succumbed to infection by D. falcata [36]. Why was this so? It would be imperative to test whether the epicortical roots emerging from D. falcata from one host can attach to a different or the same host in proximity. In a similar manner, some of the other basic questions that are intriguing in this context may be as follows: What signals enable mistletoe seeds to germinate on existing hosts? How do mistletoes expand their host tropism in light of the ever-increasing list of newer hosts? Answers to these questions would offer more useful and efficient strategies to control mistletoes. One such strategy could consider manipulating plant signaling by transgenic and epigenetic interventions, and these may easily be integrated into pest management plans such as silviculture. Both mistletoes (for infection) and their host(s) (for defense) inflict changes in in each other’s physiological and chemical profiles [53,86]. There is still uncertainty as to whether mistletoe–host interactions involve any chemical warning signals analogous to the aposematism seen in other parasites [363]. On the mistletoe’s end, some investigative leads have been gained; for example, the gelatinous seed coat material in Phoradendron spp., viscin, is known to signal host/non-host responses [87,88]. Furthermore, to initiate attachment with the host, the mistletoes require seed germination stimulants (such as ethylene and nucleophilic protein receptor) and a diversity of haustoria-inducing factors (HIFs) [275,364]. HIFs such as xyloglucan endotransgycosylases (XETs), expansins, glucanases, and cell wall hydrolases are expressed in mistletoes [89]. XETs may facilitate haustorial progression into hosts by delinking xyloglucans, which make up the secondary walls in the host vasculature [365,366]. Other HIFs comprise strigolactones, quinones, and flavonoids [275,367,368,369]. Oxidative degradation of host cell wall lignin by the action of peroxidases and H2O2 may also activate HIFs [370]. The defense signals to mistletoes from the host side are analogous to that against herbivores and other biotic stressors [9,108,371,372,373]. These are marked by anatomical variations such as lignification, suberization [116,374,375], and the formation of secondary growth-like wound periderm [292,376], all of which variously oppose progression processes from mistletoes [116]. In addition, hosts respond to mistletoe attack by making inherent changes to their hormone profiles, such as abscisic acid, jasmonic acid, salicylic acid, and indole-6-aminoacid [374]. This may increase levels of potent secondary metabolites such as tannins, terpenes, and phenolic compounds [9,61,109,130,332,372,373]. Furthermore, the infection site may display the accumulation of volatile compounds and ROS, in turn triggering enzyme cascades that signal for apoptosis, culminating in defoliation [61,130,374]. The signaling crosstalk involved during the mistletoe life cycle over a host should therefore be examined for potential candidates that drive the epigenetic control over chemical dynamics. It is also likely that a precise understanding of the epigenetic framework that governs mistletoe–insect interactions may facilitate easier and more effective screening for MBCAs.
5.9. Smart Mistletoe Management: How the 21st Century Can Mitigate the Mistletoe Problem
The literature resources do not majorly document any new developments with conventional mistletoe management approaches due to technical glitches or technological paucity. Recently, a group described a novel handheld tool, ‘Mistletoe Eradicator’, which simultaneously provides both mechanical pruning and chemical treatment [377]. The 21st century offers a huge range of possibilities with advancements into big data, automation, artificial intelligence, and platforms such as the Internet of Things (IoTs). Newer technologies such as artificial intelligence, satellite imaging (remote sensing), image recognition, drones, and weather forecasting can be utilized in substantiating smart solutions for mistletoe pest management programs (Figure 6). For example, intelligent drones may permit the aerial screening of forest canopies for mistletoes, as well as for carrying out physical (pruning) and chemical treatments (herbicide spraying, branch injections, etc.). To make this possible, detection algorithms would need precision in screening and classifying mistletoes, which might face challenges resulting from the huge diversity of mistletoes, their growth stages, geographical settings, climates, canopy structures, size, density, etc. [86,378,379,380,381,382,383]. Moreover, many mistletoe species are known to mimic their hosts in foliar structures [139]. Moser and Campione showed the successful use of Google Earth as a remote sensing tool to detect eastern spruce dwarf mistletoe in the forests of Minnesota [384]. A group from Mexico recently developed a genetic programming-based algorithm that screens mistletoe Phoradendron velutinum using multispectral aerial images collected on a radiation sensor fitted to an unmanned aerial vehicle [385]. This algorithm classifies P. velutinum based on its flowering stage. Before this, several groups had developed other algorithms based on thermal imaging, hyperspectral lines, convolutional neural networks, canopy height, colorimetry, etc. [386,387,388,389,390,391,392]. If developments into these dimensions are studied further, the effective selection of conventional and biotechnological treatment approaches may lead to a productive synchrony with silviculture. Silvicultural decisions such as treatment area and treatment schedule depend on precise knowledge of and attention to the epidemiology of mistletoes and their pathogens, mistletoe population dynamics, and the silvics of the mistletoes’ hosts [34]. In this regard, even automation and artificial intelligence solutions may offer better time-saving, labor-saving, and cost-saving prospects, as well as being amenable to modifications on a case-by-case basis.
6. Conclusions
Mistletoes have undoubtedly acquired pest status in many countries [26]. By drawing resources from other plants in communities with higher trophic levels, they parallel herbivores [108]. They are viewed as robbers by fruit and timber farmers as they present a constant problem as parasitic plants in commercial plantations. In spite of their inconsistency, cost and labor-intensiveness, a lack of technological advancements in terms of delivery systems, the impact on nearby plants, ecological disturbances, and negative environmental effects, physical and chemical control approaches have been somewhat utilized in attempting to manage this concern and have proven ineffective, if not failed. Integrated management programs, though deemed relatively effective, are practiced only on small scales and in a few countries [262]. Despite the fact that many organismic entities have been discovered to be detrimental to mistletoes, exploring and exploiting their potential as successful MBCAs has not been well realized to date. Biocontrol strategies can be synchronized with silvicultural approaches; given that models where MBCA treatment options, schedules, and mistletoe life cycles are available, this could help in framing preferred plantation plans. Other than this, where MBCAs are not a practicable solution, technological advancements are highly sought to resolve drawbacks with physical and chemical treatment methods. During our discussion, we explored the smart interventions and big data trends of 21st-century technological advancements. These ideas and opinions hold strong potential for the combination of conventional and bio/technological approaches (Figure 6). Control strategies must take into account the biological interactions and dynamics among community structures, which maintain the niche’s ecological propensities.
Considering the aforementioned promising approaches, and not neglecting the ecological value of the mistletoes (in being a keystone resource for biodiversity), the genetic engineering of hosts to develop, induce, and enhance specific resistance will be greatly rewarding. Even though its benefits include being environmentally safe and with no use of chemical agents, due to the additional labor, costs, complicated management, as well as potential to wipe out the parasite seed banks on hosts, this approach has remained unrealized [262]. Such transgenic trees would satisfy both (i) the conservation of the biodiversity and medicinal value aspects of mistletoes and (ii) aims to foresee a fruit and timber business that is unaffected by mistletoe. Successful examples of the transgenic approach are known in relation to other parasitic weeds, such as Striga, Orobanche, and Cuscuta [393,394,395,396]. In this regard, in this study, we intended to supply blueprints for transgenic and translational research into developing and commercializing stable mistletoe-resistant tree cultivars.
In this review, the authors do not underestimate the ecological significance and medicinal value of mistletoes, both in traditional and modern treatment practices. However, this article emphasizes the development and implementation of more feasible management solutions for highly damaging mistletoes that affect commercial tree plantations. The risks surely outweigh the usefulness of mistletoes in such settings, as others pointed out long ago [230]. Thus, tree decline, combined with mistletoe, cannot be overlooked in itself as a major factor negatively impacting biodiversity and commerce. Hopefully, the biotechnological and smart management approaches discussed here, if operationalized in the future, should serve as a paradigm shift in mistletoe management.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/biology11111645/s1, Table S1: List of potential MBCAs yet known for some mistletoe genera. References [397,398,399,400,401,402,403,404,405,406,407,408,409,410,411,412,413,414,415,416] are cited in the supplementary materials.
Author Contributions
Conceptualization, G.M. and M.A.G.; validation, G.M., J.K., G.B.S., and K.C.; investigation, G.M.; resources, G.M., J.K., M.P., and S.K.D.; data curation, G.M., M.A.G., M.P., and G.B.S.; writing—original draft preparation, G.M.; writing—review and editing, G.M. and M.A.G.; figures and visualization, G.M.; supervision, G.M. All authors have read and agreed to the published version of the manuscript.
Funding
The authors acknowledge the financial support for the publication of this article, received from the College of Science, United Arab Emirates University, UAE.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank the UIBT fraternity at Chandigarh University for the administrative and technical support. The institute, however, had no role in shaping the manuscript and/or in the decision to publish.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Singh, G.B.; Mudgal, G.; Vinayak, A.; Kaur, J.; Chand, K.; Parashar, M.; Verma, A.K.; Goswami, A.; Sharma, S. Molecular Communications between Plants and Microbes. In Plant-Microbial Interactions and Smart Agricultural Biotechnology; Tyagi, S., Kumar, R., Saharan, B., Nadda, A.K., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 147–184. [Google Scholar]
- Yopp, J.H.; Dickison, W.C.; Rothwell, G.W.; Lambers, H.; Schmid, R.; Woodwell, G.M. Plant. Encycl. Br. 2021. Available online: https://www.britannica.com/plant/plant (accessed on 8 October 2022).
- Satterthwaite, D.; McGranahan, G.; Tacoli, C. Urbanization and its implications for food and farming. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2010, 365, 2809–2820. [Google Scholar] [CrossRef]
- Chapagain, T.; Raizada, M.N. Impacts of natural disasters on smallholder farmers: Gaps and recommendations. Agric. Food Secur. 2017, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- FAO. The Impact of Disasters and Crises on Agriculture and Food Security: 2021; Food and Agriculture Organization: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Dubey, P.K.; Singh, G.S.; Abhilash, P.C. Agriculture in a changing climate. In Adaptive Agricultural Practices; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–10. [Google Scholar]
- Oshunsanya, S.O.; Nwosu, N.J.; Li, Y. Abiotic stress in agricultural crops under climatic conditions. In Sustainable Agriculture, Forest and Environmental Management; Springer: Singapore, 2019; pp. 71–100. [Google Scholar]
- Joos, L.; De Tender, C. Soil under stress: The importance of soil life and how it is influenced by (micro)plastic pollution. Comput. Struct. Biotechnol. J. 2022, 20, 1554–1566. [Google Scholar] [CrossRef] [PubMed]
- Klutsch, J.G.; Erbilgin, N. Dwarf mistletoe infection in jack pine alters growth-defense relationships. Tree Physiol. 2018, 38, 1538–1547. [Google Scholar] [CrossRef] [PubMed]
- Ehleringer, J.; Schulze, E.-D.; Ziegler, H.; Lange, O.; Farquhar, G.; Cowar, I. Xylem-tapping mistletoes: Water or nutrient parasites? Science 1985, 227, 1479–1481. [Google Scholar] [CrossRef] [PubMed]
- Schulze, E.D.; Turner, N.; Glatzel, G. Carbon, water and nutrient relations of two mistletoes and their hosts: A hypothesis. Plant Cell Environ. 1984, 7, 293–299. [Google Scholar] [CrossRef]
- The Editors of Encyclopaedia Britannica. Mistletoe. Encyclopedia Britannica. Available online: https://www.britannica.com/plant/mistletoe (accessed on 27 October 2022).
- Halloin, L. Dwarf Mistletoe Biology and Management in Southeast Region; Washington State Department of Natural Resources: Washington, DC, USA, 2003; pp. 1–8. Available online: https://www.dnr.wa.gov/Publications/rp_fh_wadnrdwarfmistletoe.pdf (accessed on 27 October 2022).
- MCFB. Dwarf Mistletoe of Conifers. Available online: https://www.gov.mb.ca/nrnd/forest/pubs/forest_lands/health/dwarf_mistletoe_brochure.pdf (accessed on 22 October 2021).
- Ferguson, B.A. State Forester Forum-Dwarf Mistletoes; Ferguson Forest Pathology Consulting, Inc.: Missoula, MT, USA, 2014; p. 9. Available online: https://www.idl.idaho.gov/wp-content/uploads/sites/2/forestry/forester-forums/id24.pdf (accessed on 8 October 2022).
- Mallams, K.M.; Mathiasen, R.L. Mistletoes on Hardwoods in the United States. For. Insect Dis. Leafl. 2010, 147, 1–10. [Google Scholar]
- Aly, R. Conventional and biotechnological approaches for control of parasitic weeds. Vitr. Cell. Dev. Biol. Plant 2007, 43, 304–317. [Google Scholar] [CrossRef]
- Nickrent, D.L. Parasitic angiosperms: How often and how many? Taxon 2020, 69, 5–27. [Google Scholar] [CrossRef]
- Parker, C. Protection of crops against parasitic weeds. Crop Prot. 1991, 10, 6–22. [Google Scholar] [CrossRef]
- Kaiser, B.; Vogg, G.; Fürst, U.B.; Albert, M. Parasitic plants of the genus Cuscuta and their interaction with susceptible and resistant host plants. Front. Plant Sci. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauerborn, J.; Müller-Stöver, D.; Hershenhorn, J. The role of biological control in managing parasitic weeds. Crop Prot. 2007, 26, 246–254. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M. Innovations in parasitic weeds management in legume crops. A review. Agron. Sustain. Dev. 2012, 32, 433–449. [Google Scholar] [CrossRef]
- Albert, M.; Axtell, M.J.; Timko, M.P. Mechanisms of resistance and virulence in parasitic plant–host interactions. Plant Physiol. 2020, 185, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Aparicio, M.; Delavault, P.; Timko, M.P. Management of Infection by Parasitic Weeds: A Review. Plants 2020, 9, 1184. [Google Scholar] [CrossRef] [PubMed]
- Coleman, E. Menace of the mistletoe. Vic. Nat. 1949, 66, 24–32. [Google Scholar]
- Watson, D.M.; Cook, M.E.; Fadini, R.R. Towards best-practice management of mistletoes in horticulture. Botany 2020, 98, 489–498. [Google Scholar] [CrossRef]
- Geils, B.W. Mistletoes of North American Conifers; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2002. [Google Scholar] [CrossRef]
- Pearson, G.; Marsh, R. Timber Growing and Logging Practice in the Southwest and in the Black Hills Region. Available online: https://naldc.nal.usda.gov/download/CAT86200474/PDF (accessed on 22 October 2022).
- Reid, N.; Shamoun, S.F. Contrasting research approaches to managing mistletoes in commercial forests and wooded pastures. Botany 2008, 87, 1–9. [Google Scholar] [CrossRef]
- Watson, D.M. Mistletoe—A Keystone Resource in Forests and Woodlands Worldwide. Annu. Rev. Ecol. Syst. 2001, 32, 219–249. [Google Scholar] [CrossRef] [Green Version]
- Madisa, M.E.; Assefa, Y.; Kelemoge, O.D.; Mathowa, T.; Segwagwe, A. Incidence and level of mistletoe infestation in tree species at Botswana University of Agriculture and Natural Resources’ Sebele Content Farm Campus, Botswana. Int. J. Environ. Agric. Res. 2017, 3, 53–58. [Google Scholar] [CrossRef]
- Stinziano, J. Mistletoe-ing around the Tree Reduces Growth in Forests. Available online: https://botany.one/2017/12/mistletoe-ing-around-tree-reduces-growth-forests/ (accessed on 22 October 2022).
- Rist, L.; Shaanker, R.U.; Ghazoul, J. The Spatial Distribution of Mistletoe in a Southern Indian Tropical Forest at Multiple Scales. Biotropica 2011, 43, 50–57. [Google Scholar] [CrossRef]
- Shamoun, S.; DeWald, L. Management strategies for dwarf mistletoes: Biological, chemical, and genetic approaches. In Mistletoes of North American Conifers; Geils, B.W., Tovar, J.C., Moody, B., Eds.; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Washington, DC, USA, 2002; Volume 98, pp. 75–82. [Google Scholar]
- Choi, S.-U.; Kim, S.T.; Han, D.G.; Hwang, Y.-H.; Lee, K.Y.; Kim, D.U.; Cho, K.H.; Park, S.Y.; Kim, H.-C.; Kim, S.-B.; et al. Comparative Assessment of Biological Activities of Mistletoes for Cosmetic Applications: Viscum Album Var. Coloratum (Kom.) Ohwi and Loranthus Tanakae Franch. & Sav. J. Cosmet. Sci. 2019, 70, 235–245. [Google Scholar] [PubMed]
- Mudgal, G.; Mudgal, B. Evidence for unusual choice of host and haustoria by Dendrophthoe falcata (Lf) Ettingsh, a leafy mistletoe. Arch. Phytopathol. Plant Prot. 2011, 44, 186–190. [Google Scholar] [CrossRef]
- Mudgal, G.; Mudgal, B.; Gururani, M.A.; Jelli, V. Pseudaulacaspis cockerelli (Cooley) hyperparasitizing Dendrophthoe falcata (Lf) Ettingsh. Arch. Phytopathol. Plant Prot. 2011, 44, 282–286. [Google Scholar] [CrossRef]
- Singh, S.R.; Piloo, N.; Senjam, P.; Hemanta, L. Need of awareness programme to control the loranthus weed–Helixanthera ligustrina. Agri-India Today 2021, 1, 10–13. [Google Scholar]
- Arce-Acosta, I.; Suzán-Azpiri, H.; García-Rubio, O. Biotic factors associated with the spatial distribution of the mistletoe Psittacanthus calyculatus in a tropical deciduous forest of central Mexico. Bot. Sci. 2016, 94, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Fadini, R.F.; Fischer, E.; Castro, S.J.; Araujo, A.C.; Ornelas, J.F.; de Souza, P.R. Bat and bee pollination in Psittacanthus mistletoes, a genus regarded as exclusively hummingbird-pollinated. Ecology 2018, 99, 1239–1241. [Google Scholar] [CrossRef]
- Infante, S.D.; Lara, C.; del Coro Arizmendi, M.; Eguiarte, L.E.; Ornelas, J.F. Reproductive ecology and isolation of Psittacanthus calyculatus and P. auriculatus mistletoes (Loranthaceae). PeerJ 2016, 4, e2491. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Crespo, M.J.; Lara, C.; Ornelas, J.F. Uncorrelated mistletoe infection patterns and mating success with local host specialization in Psittacanthus calyculatus (Loranthaceae). Evol. Ecol. 2016, 30, 1061–1080. [Google Scholar] [CrossRef]
- Teodoro, G.S.; van den Berg, E.; Santos, M.d.C.N.; de Freitas Coelho, F. How does a Psittacanthus robustus Mart. population structure relate to a Vochysia thyrsoidea Pohl. host population? Flora-Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 797–801. [Google Scholar] [CrossRef]
- Aliero, B.; Samaila, A. The occurrence of Parasitic mistletoe (Tapinanthus spp) on Parkia biglobosa (Jacq.) Benth (African Locust Bean Tree) in Yauri Local Government Area, Kebbi State. Niger. J. Basic Appl. Sci. 2000, 9, 5–10. [Google Scholar]
- Edagbo, D.E.; Ajiboye, T.O.; Borokini, T.I.; Ighere, D.A.; Alowonle, A.A.; Clement, M. The Influence of African Mistletoe (Tapinanthus bangwensis) on the Conservation Status and Productivity of Irvingia gabonensis in Moor Plantation Area of Ibadan, Nigeria. Int. J. Curr. Res. 2012, 4, 484–487. [Google Scholar] [CrossRef]
- Kwon-Ndung, E.; Ismaila, A. Prospects of host resistance in improved and domesticated species of Parkia biglobosa to African mistletoes (Tapinanthus spp.) in Central Nigeria. Electron. J. Environ. Agric. Food Chem. 2009, 8, 382–388. [Google Scholar]
- Ávalos, V.M.C.; Collazo, I.V.; Flores, H.J.M.; Castillo, J.V. Impacto de tierra de diatomeas sobre Arceuthobium globosum Hawksworth & Wiens subsp. grandicaule en Pinus pseudostrobus Lindl. Rev. Mex. Cienc. For. 2010, 1, 39–46. [Google Scholar]
- Gilbert, J.A. The Biology of Dwarf Mistletoes (Arceuthobium spp.) in Manitoba. Master Thesis, University of Manitoba, Winnipeg, MB, Canada, 1984. [Google Scholar]
- Hernández-Benítez, R.; Cano-Santana, Z.; Castellanos-Vargas, I. Incidencia de infestación de Arceuthobium globosum grandicaule (Hawks. y Wiens) en Pinus hartwegii (Lindl.). Cienc. For. En México 2005, 30, 79–86. [Google Scholar]
- Queijeiro-Bolanos, M.E.; Cano-Santana, Z.; Castellanos-Vargas, I. Does disturbance determine the prevalence of dwarf mistletoe (Arceuthobium, Santalales: Viscaceae) in Central Mexico? Rev. Chil. De Hist. Nat. 2013, 86, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Shaw, D.C.; Agne, M.C.J.B. Fire and dwarf mistletoe (Viscaceae: Arceuthobium species) in western North America: Contrasting Arceuthobium tsugense and Arceuthobium americanum. Botany 2017, 95, 231–246. [Google Scholar] [CrossRef]
- Villier, J.A.d.; Reblin, J.S.; Logan, B.A. Needle properties of host white spruce (Picea glauca [Moench] Voss) experiencing eastern dwarf mistletoe (Arceuthobium pusillum Peck) infections of differing severity. Botany 2017, 95, 295–305. [Google Scholar] [CrossRef]
- Anselmo-Moreira, F.; Teixeira-Costa, L.; Ceccantini, G.; Furlan, C.M. Mistletoe effects on the host tree Tapirira guianensis: Insights from primary and secondary metabolites. Chemoecology 2019, 29, 11–24. [Google Scholar] [CrossRef]
- Solís-Gracia, V.; Suzán-Azpiri, H. Análisis de la distribución espacial del muérdago (Phoradendron californicum) en el sur del Desierto Sonorense. Cactáceas Suculentas Mex. 2014, 59, 11–28. [Google Scholar]
- Wiens, D.; Hawksworth, F. New species of Phoradendron (Viscaceae) from Mexico and Guatemala and a synopsis of species in section Pauciflorae. Aliso J. Syst. Evol. Bot. 2002, 21, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Mir, N.; Sultan, S. White-berry mistletoe (Viscum album L.): A hemiparasitic plant: Occurrence and ethnobotanical use in Kashmir. J. Pharmacogn. Phytochem. 2018, 7, 1831–1833. [Google Scholar]
- Alam, M. A critical review on the biology and control of Loranthaceae with a particular reference to Bangladesh. Bano Biggyan Patrika 1984, 13, 7–18. [Google Scholar]
- Barbu, C. Impact of mistletoe attack (Viscum album ssp. abietis) on the radial growth of silver fir. A case study in the North of Eastern Carpathians. Ann. For. Res. 2009, 52, 89–96. [Google Scholar] [CrossRef]
- Bilgili, E.; Coskuner, K.A.; Öztürk, M. Leaf area–sapwood area relationship in Scots pine (Pinus sylvestris L.) under mistletoe (Viscum album ssp. austriacum) infection. Dendrobiology 2020, 84, 1–11. [Google Scholar] [CrossRef]
- Dobbertin, M.; Rigling, A. Pine mistletoe (Viscum album ssp. austriacum) contributes to Scots pine (Pinus sylvestris) mortality in the Rhone valley of Switzerland. For. Pathol. 2006, 36, 309–322. [Google Scholar] [CrossRef]
- Mutlu, S.; Osma, E.; Ilhan, V.; Turkoglu, H.I.; Atici, O. Mistletoe (Viscum album) reduces the growth of the Scots pine by accumulating essential nutrient elements in its structure as a trap. Trees 2016, 30, 815–824. [Google Scholar] [CrossRef]
- Ozturk, M.; Coskuner, K.A.; Serdar, B.; Atar, F.; Bilgili, E. Impact of white mistletoe (Viscum album ssp. abietis) infection severity on morphology, anatomy and photosynthetic pigment content of the needles of cilicican fir (Abies cilicica). Flora 2022, 294, 152135. [Google Scholar] [CrossRef]
- Ozturk, M.; Coskuner, K.A.; Usta, Y.; Serdar, B.; Bilgili, E. The effect of mistletoe (Viscum album) on branch wood and needle anatomy of Scots pine (Pinus sylvestris). IAWA J. 2019, 40, 352–365. [Google Scholar] [CrossRef]
- Glatzel, G.; Geils, B. Mistletoe ecophysiology: Host–parasite interactions. Botany 2008, 87, 10–15. [Google Scholar] [CrossRef]
- Calvin, C.L.; Wilson, C.A. Comparative morphology of epicortical roots in Old and New World Loranthaceae with reference to root types, origin, patterns of longitudinal extension and potential for clonal growth. Flora-Morphol. Distrib. Funct. Ecol. Plants 2006, 201, 51–64. [Google Scholar] [CrossRef]
- Barlow, B. MIstletoes in Australia. Available online: http://www.anbg.gov.au/mistletoe/haustoria.html (accessed on 12 January 2020).
- Wilson, C.A.; Calvin, C.L. An origin of aerial branch parasitism in the mistletoe family, Loranthaceae. Am. J. Bot. 2006, 93, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Stewart, G.R. Effect of Potassium Levels on the Stomatal Behavior of the Hemi-Parasite Striga hermonthica. Plant Physiol. 1990, 94, 1472–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Gullo, M.A.; Glatzel, G.; Devkota, M.; Raimondo, F.; Trifilò, P.; Richter, H. Mistletoes and mutant albino shoots on woody plants as mineral nutrient traps. Ann. Bot. 2012, 109, 1101–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunaichamy, K.S.T.K.; Paliwal, K.; Arp, P.A. Biomass and nutrient dynamics of mistletoe (Dendrophthoe falcata) and neem (Azadirachta indica) seedlings. Curr. Sci. 1999, 76, 840–842. [Google Scholar]
- Zweifel, R.; Bangerter, S.; Rigling, A.; Sterck, F.J. Pine and mistletoes: How to live with a leak in the water flow and storage system? J. Exp. Bot. 2012, 63, 2565–2578. [Google Scholar] [CrossRef] [Green Version]
- Urban, J.; Gebauer, R.; Nadezhdina, N.; Čermák, J. Transpiration and stomatal conductance of mistletoe (Loranthus europaeus) and its host plant, downy oak (Quercus pubescens). Biologia 2012, 67, 917–926. [Google Scholar] [CrossRef] [Green Version]
- Glatzel, G. Mineral nutrition and water relations of hemiparasitic mistletoes: A question of partitioning. Experiments with Loranthus europaeus on Quercus petraea and Quercus robur. Oecologia 1983, 56, 193–201. [Google Scholar] [CrossRef]
- Yang, D.; Goldstein, G.; Wang, M.; Zhang, W.-W.; Wang, A.-Y.; Liu, Y.-Y.; Hao, G.-Y. Microenvironment in the canopy rivals the host tree water status in controlling sap flow of a mistletoe species. Tree Physiol. 2017, 37, 501–510. [Google Scholar] [CrossRef]
- Rigling, A.; Eilmann, B.; Koechli, R.; Dobbertin, M. Mistletoe-induced crown degradation in Scots pine in a xeric environment. Tree Physiol. 2010, 30, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Dobbertin, M.; Hilker, N.; Rebetez, M.; Zimmermann, N.E.; Wohlgemuth, T.; Rigling, A. The upward shift in altitude of pine mistletoe (Viscum album ssp. austriacum) in Switzerland—The result of climate warming? Int. J. Biometeorol. 2005, 50, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doležal, J.; Lehečková, E.; Sohar, K.; Altman, J. Oak decline induced by mistletoe, competition and climate change: A case study from central Europe. Preslia 2016, 88, 323–346. [Google Scholar]
- Matula, R.; Svátek, M.; Pálková, M.; Volařík, D.; Vrška, T. Mistletoe infection in an oak forest is influenced by competition and host size. PLoS ONE 2015, 10, e0127055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aukema, J.E.; Martinez del Rio, C. Mistletoes as parasites and seed-dispersing birds as disease vectors: Current understanding, challenges, and opportunities. In Seed Dispersal and Frugivory: Ecology, Evolution, and Conservation; CABI International: Wallingford, UK, 2002; pp. 99–110. [Google Scholar]
- Aukema, J.E.; Martínez del Rio, C. Where does a fruit-eating bird deposit mistletoe seeds? Seed deposition patterns and an experiment. Ecology 2002, 83, 3489–3496. [Google Scholar] [CrossRef]
- Reid, N. Coevolution of mistletoes and frugivorous birds? Aust. J. Ecol. 1991, 16, 457–469. [Google Scholar] [CrossRef]
- Green, A.K.; Ward, D.; Griffiths, M.E. Directed dispersal of mistletoe (Plicosepalus acaciae) by Yellow-vented Bulbuls (Pycnonotus xanthopygos). J. Ornithol. 2009, 150, 167–173. [Google Scholar] [CrossRef]
- Mathiasen, R.L.; Nickrent, D.L.; Shaw, D.C.; Watson, D.M. Mistletoes: Pathology, systematics, ecology, and management. Plant Dis. 2008, 92, 988–1006. [Google Scholar] [CrossRef] [Green Version]
- Watson, D.M. The Relative Contribution of Specialists and Generalists to Mistletoe Dispersal: Insights from a Neotropical Rain Forest. Biotropica 2013, 45, 195–202. [Google Scholar] [CrossRef]
- Albert, S.; Rhumeur, A.; Rivière, J.L.; Chauvrat, A.; Sauroy-Toucouère, S.; Martos, F.; Strasberg, D. Rediscovery of the mistletoe Bakerella hoyifolia subsp. bojeri (Loranthaceae) on Reunion Island: Population status assessment for its conservation. Bot. Lett. 2017, 164, 229–236. [Google Scholar] [CrossRef]
- Medel, R.; Vergara, E.; Silva, A.; Kalin-Arroyo, M. Effects of vector behavior and host resistance on mistletoe aggregation. Ecology 2004, 85, 120–126. [Google Scholar] [CrossRef]
- Gill, L.S.; Hawksworth, F.G. The Mistletoes: A Literature Review; US Department of Agriculture: Washington, DC, USA, 1961. [Google Scholar]
- Berry, A.M.; Lichter, J.M.; Reid, M.S. New Methods for Mistletoe Control. Available online: http://slosson.ucdavis.edu/newsletters/Berry_199129111.pdf (accessed on 29 December 2019).
- Ko, S.M.; Kwon, Y.K.; Kim, J.H.; Song, I.-J.; Lee, H.-Y.; Choi, D.-W.; Liu, J.R.; Kim, S.W. Transcriptome analysis of mistletoe (Viscum album) haustorium development. Hortic. Environ. Biotechnol. 2014, 55, 352–361. [Google Scholar] [CrossRef]
- Scharpf, R.F.; Smith, R.S.; Vogler, D. Management of Western Dwarf Mistletoe in Ponderosa and Jeffrey Pines in Forest Recreation Areas; Pacific Southwest Research Station, Forest Service; US Department of Agriculture: Washington, DC, USA, 1988; Volume 103, 11p. [Google Scholar]
- Knutson, D. Physiology of mistletoe parasitism and disease responses in the host. In The Biology of Mistletoes; Academic Press: Sydney, Australia, 1983; pp. 295–316. [Google Scholar]
- Nickrent, D.L. Santalales (Mistletoe). In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: New York, NY, USA, 2002. [Google Scholar]
- Geils, B.; Hawksworth, F. Damage, Effects, and Importance of Dwarf Mistletoes; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2002; pp. 57–65. [Google Scholar]
- Hawksworth, F.G. Mistletoes on Introduced Trees of the World; US Department of Agriculture Forest Service: Washington, DC, USA, 1974. [Google Scholar]
- Barney, C.W.; Hawksworth, F.; Geils, B. Hosts of Viscum album. J. For. Pathol. 1998, 28, 187–208. [Google Scholar] [CrossRef]
- Hawksworth, F.G.; Wiens, D.; Geils, B.W. Arceuthobium in North America. In General Technical Report; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Washington, DC, USA, 2002; Chapter 4; pp. 29–56. [Google Scholar]
- Ritter, S.M.; Hoffman, C.M.; Ex, S.A.; Stewart, J.E. Impacts of lodgepole pine dwarf mistletoe (Arceuthobium americanum) infestation on stand structure and fuel load in lodgepole pine dominated forests in central Colorado. Botany 2017, 95, 307–321. [Google Scholar] [CrossRef]
- Shaw, D.C.; Watson, D.M.; Mathiasen, R.L. Comparison of dwarf mistletoes (Arceuthobium spp., Viscaceae) in the western United States with mistletoes (Amyema spp., Loranthaceae) in Australia—Ecological analogs and reciprocal models for ecosystem management. Aust. J. Bot. 2004, 52, 481–498. [Google Scholar] [CrossRef] [Green Version]
- Swanson, M.E.; Shaw, D.C.; Marosi, T.K. Distribution of Western Hemlock Dwarf Mistletoe (Arceuthobioum tsugense [Rosendahl] GN Jones Subsp. tsugense) in Mature and Old-growth Douglas-fir (Pseudotsuga menziesii [Mirb.] Franco) Forests. Northwest Sci. 2006, 80, 207. [Google Scholar]
- Yi, J.-S.; Kim, C.-W.; Lee, J.-K.; Meng, F.-J. Identification of Populations of Korean Mistletoe (Viscum Album L. Var. Coloratum) from Gangwon-Do in Korea by AFLP. Biotechnol. Biotechnol. Equip. 2013, 27, 4091–4097. [Google Scholar] [CrossRef]
- Kim, C.-S.; Kim, S.-Y.; Sun, B.-Y.; Yi, J.S. A review of the taxonomic and ecological characteristics of Korean mistletoe types (Viscum, Korthalsella, Loranthus and Taxillus). Korean J. Plant Taxon. 2013, 43, 81–89. [Google Scholar] [CrossRef]
- Sivaramakrishna, P.; Yugandhar, P.; Ekka, G.A. A new species Dendrophthoe laljii (Loranthaceae) infesting Artocarpus heterophyllus Lam. (Moraceae) in Andaman and Nicobar Islands, India. J. Asia-Pac. Biodivers. 2021, 14, 452–459. [Google Scholar] [CrossRef]
- Singh, L.J.; Ranjan, V.; Rasingam, L.; Swamy, J. A new species of genus Dendrophthoe Mart. (Lorantheae-Loranthaceae) from the Peninsular India. J. Asia-Pac. Biodivers. 2020, 13, 487–493. [Google Scholar] [CrossRef]
- Devkota, M.P. Biology of mistletoes and their status in Nepal Himalaya. Himal. J. Sci. 2005, 3, 84–88. [Google Scholar] [CrossRef]
- Bilgili, E.; Coskuner, K.A.; Baysal, I.; Ozturk, M.; Usta, Y.; Eroglu, M.; Norton, D. The distribution of pine mistletoe (Viscum album ssp. austriacum) in Scots pine (Pinus sylvestris) forests: From stand to tree level. Scand. J. For. Res. 2020, 35, 20–28. [Google Scholar] [CrossRef]
- Aukema, J.E. Vectors, viscin, and Viscaceae: Mistletoes as parasites, mutualists, and resources. Front. Ecol. Environ. 2003, 1, 212–219. [Google Scholar] [CrossRef]
- Kuijt, J. Dwarf mistletoes. Bot. Rev. 1955, 21, 569–619. [Google Scholar] [CrossRef]
- Pennings, S.C.; Callaway, R.M. Parasitic plants: Parallels and contrasts with herbivores. Oecologia 2002, 131, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Press, M.C.; Phoenix, G.K. Impacts of parasitic plants on natural communities. New Phytol. 2005, 166, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Okubamichael, D.Y.; Griffiths, M.E.; Ward, D. Host specificity, nutrient and water dynamics of the mistletoe Viscum rotundifolium and its potential host species in the Kalahari of South Africa. J. Arid Environ. 2011, 75, 898–902. [Google Scholar] [CrossRef]
- Yan, C.-F.; Gessler, A.; Rigling, A.; Dobbertin, M.; Han, X.-G.; Li, M.-H. Effects of mistletoe removal on growth, N and C reserves, and carbon and oxygen isotope composition in Scots pine hosts. Tree Physiol. 2016, 36, 562–575. [Google Scholar] [CrossRef] [Green Version]
- Schulze, E.-D.; Kelliher, F.M.; Körner, C.; Lloyd, J.; Leuning, R. Relationships among maximum stomatal conductance, ecosystem surface conductance, carbon assimilation rate, and plant nitrogen nutrition: A global ecology scaling exercise. Annu. Rev. Ecol. Syst. 1994, 25, 629–662. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Cook, C.S.; Tieszen, L.L. Comparative water use and nitrogen relationships in a mistletoe and its host. Oecologia 1986, 68, 279–284. [Google Scholar] [CrossRef]
- Sangüesa-Barreda, G.; Linares, J.C.; Camarero, J.J. Mistletoe effects on Scots pine decline following drought events: Insights from within-tree spatial patterns, growth and carbohydrates. Tree Physiol. 2012, 32, 585–598. [Google Scholar] [CrossRef] [Green Version]
- Scalon, M.C.; dos Reis, S.A.; Rossatto, D.R. Shifting from acquisitive to conservative: The effects of Phoradendron affine (Santalaceae) infection in leaf morpho-physiological traits of a Neotropical tree species. Aust. J. Bot. 2017, 65, 31–37. [Google Scholar] [CrossRef]
- Teixeira-Costa, L.; Ceccantini, G. Embolism increase and anatomical modifications caused by a parasitic plant: Phoradendron crassifolium (Santalaceae) on Tapirira guianensis (Anacardiaceae). IAWA J. 2015, 36, 138–151. [Google Scholar] [CrossRef]
- Roth, L.F. Dwarf mistletoe-induced mortality in northwest ponderosa pine growing stock. West. J. Appl. For. 2001, 16, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Raftoyannis, Y.; Radoglou, K.; Bredemeier, M. Effects of mistletoe infestation on the decline and mortality of Abies cephalonica in Greece. Ann. For. Res. 2015, 58, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Fisher, J.T. Water relations of mistletoes and their hosts. In The Biology of Mistletoes; Calder, M., Bernhardt, P., Eds.; Academic: Sydney, Australia, 1983; pp. 161–184. [Google Scholar]
- Parker, C.; Riches, C.R. Parasitic Weeds of the World: Biology and Control; CAB International: Waringford, UK, 1993; Volume 48. [Google Scholar]
- Szmidla, H.; Tkaczyk, M.; Plewa, R.; Tarwacki, G.; Sierota, Z. Impact of Common Mistletoe (Viscum album L.) on Scots Pine Forests—A Call for Action. Forests 2019, 10, 847. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, S.M.; Kartoolinejad, D.; Mirnia, S.K.; Tabibzadeh, Z.; Akbarinia, M.; Shayanmehr, F. The effects of Viscum album L. on foliar weight and nutrients content of host trees in Caspian forests (Iran). Pol. J. Ecol. 2007, 55, 579. [Google Scholar]
- Catal, Y.; Carus, S. Effect of pine mistletoe on radial growth of crimean pine (Pinus nigra) in Turkey. J. Environ. Biol. 2011, 32, 263–270. [Google Scholar]
- Queijeiro-Bolaños, M.E.; Malda-Barrera, G.X.; Carrillo-Angeles, I.G.; Suzán-Azpiri, H. Contrasting gas exchange effects on the interactions of two mistletoe species and their host Acacia schaffneri. J. Arid Environ. 2020, 173, 104041. [Google Scholar] [CrossRef]
- Bilgili, E.; Ozturk, M.; Coskuner, K.; Baysal, I.; Serdar, B.; Yavuz, H.; Eroglu, M.; Usta, Y. Quantifying the effect of pine mistletoe on the growth of Scots pine. For. Pathol. 2018, 48, e12435. [Google Scholar] [CrossRef]
- Kollas, C.; Gutsch, M.; Hommel, R.; Lasch-Born, P.; Suckow, F. Mistletoe-induced growth reductions at the forest stand scale. Tree Physiol. 2017, 38, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Pilichowski, S.; Filip, R.; Koscielska, A.; Zaroffe, G.; Zyzniewska, A.; Iszkulo, G. Influence of Viscum album ssp austriacum (Wiesb.) Vollm. on tree radial growth of Pinus sylvestris L. Sylwan 2018, 162, 452–459. [Google Scholar]
- Hawksworth, F.G.; Wiens, D. Dwarf Mistletoes: Biology, Pathology, and Systematics; Diane Publishing: Washington, DC, USA; Department of Agriculture, Forest Service: Washington, DC, USA, 1996; p. 410. [Google Scholar]
- Varga, I.; Taller, J.; Baltazár, T.; Hyvönen, J.; Poczai, P. Leaf-spot disease on European mistletoe (Viscum album) caused by Phaeobotryosphaeria visci: A potential candidate for biological control. Biotechnol. Lett. 2012, 34, 1059–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lázaro-González, A.; Hódar, J.A.; Zamora, R. Mistletoe Versus Host Pine: Does Increased Parasite Load Alter the Host Chemical Profile? J. Chem. Ecol. 2019, 45, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Lázaro-González, A.; Gargallo-Garriga, A.; Hódar, J.A.; Sardans, J.; Oravec, M.; Urban, O.; Peñuelas, J.; Zamora, R. Implications of mistletoe parasitism for the host metabolome: A new plant identity in the forest canopy. Plant. Cell Environ. 2021, 44, 3655–3666. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.C.; Guimarães, A.F.; Teodoro, G.S.; Bastos, S.S.; de Castro, E.M.; van den Berg, E. The enemy within: The effects of mistletoe parasitism on infected and uninfected host branches. Plant Ecol. 2021, 222, 639–645. [Google Scholar] [CrossRef]
- Gehring, C.A.; Whitham, T.G. Reduced mycorrhizae on Juniperus monosperma with mistletoe: The influence of environmental stress and tree gender on a plant parasite and a plant-fungal mutualism. Oecologia 1992, 89, 298–303. [Google Scholar] [CrossRef]
- Cullings, K.; Raleigh, C.; Vogler, D.R. Effects of severe dwarf mistletoe infection on the ectomycorrhizal community of a Pinus contorta stand in Yellowstone Park. Botany 2005, 83, 1174–1180. [Google Scholar] [CrossRef]
- Stermitz, F.R.; Belofsky, G.N.; Ng, D.; Singer, M.C. Quinolizidine alkaloids obtained by Pedicularis semibarbata (Scrophulariaceae) from Lupinus fulcratus (Leguminosae) fail to influence the specialist herbivore Euphydryas editha (Lepidoptera). J. Chem. Ecol. 1989, 15, 2521–2530. [Google Scholar] [CrossRef]
- Marko, M.D.; Stermitz, F.R. Transfer of alkaloids from Delphinium to Castilleja via root parasitism. Norditerpenoid alkaloid analysis by electrospray mass spectrometry. Biochem. Syst. Ecol. 1997, 25, 279–285. [Google Scholar] [CrossRef]
- Schneider, M.J.; Stermitz, F.R. Uptake of host plant alkaloids by root parasitic Pedicularis species. Phytochemistry 1990, 29, 1811–1814. [Google Scholar] [CrossRef]
- Ehleringer, J.; Ullmann, I.; Lange, O.; Farquhar, G.; Cowan, I.; Schulze, E.-D.; Ziegler, H. Mistletoes: A hypothesis concerning morphological and chemical avoidance of herbivory. Oecologia 1986, 70, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Barlow, B.A.; Wiens, D. Host-Parasite Resemblance in Australian Mistletoes: The Case for Cryptic Mimicry. Evolution 1977, 31, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.D.; Mescher, M.C.; De Moraes, C.M. Implications of bioactive solute transfer from hosts to parasitic plants. Curr. Opin. Plant Biol. 2013, 16, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Blick, R.A.J.; Burns, K.C.; Moles, A.T. Predicting network topology of mistletoe–host interactions: Do mistletoes really mimic their hosts? Oikos 2012, 121, 761–771. [Google Scholar] [CrossRef]
- Beatty, J.S.; Mathiasen, R.L. Dwarf Mistletoes of Ponderosa Pine; US Department of Agriculture, Forest Service: Washington, DC, USA, 2003. [Google Scholar]
- Knutson, D. Biological and chemical control of dwarf mistletoe. In Proceedings of the Symposium on Dwarf Mistletoe Control through Forest Management, Berkeley, CA, USA, 11–13 April 1978; Gen. Tech. Rep. PSW-31. Scharpf, R.F., Parmeter, J.R., Jr., Eds.; U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1978; pp. 151–155. [Google Scholar]
- Milenkovic, M.; Karadzic, D.; Janjic, V.; Mihajlovic, L. Possibilities of mistletoe control. In Proceedings of the Peti Kongres o Korovima, Banja Koviljaca, Serbia, 18–21 June 1996. [Google Scholar]
- Watson, D. Reconnaissance and Recommendations for Mistletoe Management in Macadamia Orchards; Final Report; Hort Innovation: North Sydney, Australia, 2019; pp. 1–30. [Google Scholar]
- Walldén, B. Misteln vid dess nordgräns: Die Mistel an Ihrer Nordgrenze. Svensk Botanisk Tidskrift 1961, 55, 123. [Google Scholar]
- Weber, H. Untersuchungen zur Entwicklungsweise der Laubholzmistel Viscum album L.(Viscaceae) und über Zuwachsraten während ihrer ersten Stadien. Beiträge Zur Biol. Der Pflanz. 1993, 67, 319–331. [Google Scholar]
- Hoffman, J. Management Guide for Dwarf Mistletoe. Arceuthobium spp. Forest Health Protection and State Forestry Organizations. Available online: https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5187427.pdf (accessed on 22 October 2022).
- Zielke, K.; Bancroft, B. Introduction to Silvicultural Systems (Web-Based Workbook). Available online: https://www.for.gov.bc.ca/hfd/pubs/ssintroworkbook/append2.htm (accessed on 20 July 2020).
- Parmeter, J.; Scharpf, R.F. Spread of Dwarf Mistletoe from Discrete Seed Sources into Young Stands of Ponderosa and Jeffrey Pines; US Forest Service: Washington, DC, USA; US Department of Agriculture, Pacific Southwest Forest and Range: Vallejo, CA, USA, 1972; Volume 269. [Google Scholar]
- Scharpf, R.F.; Parmeter, J. Seed Production and Dispersal by Dwarf Mistletoe in Overstory Jeffrey Pines in California; US Forest Service: Washington, DC, USA; US Department of Agriculture, Pacific Southwest Forest and Range: Berkeley CA, USA, 1971; Volume 247. [Google Scholar]
- Scharpf, R.F.; Parmeter, J.R., Jr. Spread of Dwarf Mistletoe into Jeffrey Pine Plantation-Trees Infected after 22 Years; U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1976; 6p. [Google Scholar]
- Reid, N.; Yan, Z. Mistletoes and Other Phanerogams Parasitic on Eucalypts; CSIRO: Canberra, Australia, 2000; pp. 353–384. [Google Scholar]
- Kelly, P.; Reid, N.; Davies, I. Effects of experimental burning, defoliation, and pruning on survival and vegetative resprouting in mistletoes (Amyema miquelii and Amyema pendula). Int. J. Plant Sci. 1997, 158, 856–861. [Google Scholar] [CrossRef]
- Conklin, D.A.; Geils, B.W. Survival and sanitation of dwarf mistletoe-infected ponderosa pine following prescribed underburning. West. J. Appl. For. 2008, 23, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Koonce, A.L.; Roth, L.F. The effects of prescribed burning on dwarf mistletoe in ponderosa pine. In Proceedings of the Sixth Conference on Fire and Forest Meteorology, Seattle, WA, USA, 22–24 April 1980; pp. 22–24. [Google Scholar]
- Ritter, S.; Hoffman, C.; Stewart, J.; Zimmerman, T. The influence of prescribed crown fire on lodgepole pine dwarf mistletoe (Arceuthobium americanum) populations 33 years post-fire. For. Pathol. 2018, 48, e12419. [Google Scholar] [CrossRef]
- Alexander, M.E.; Hawksworth, F.G. Fire and dwarf mistletoes in North American coniferous forests. J. For. 1976, 74, 446–449. [Google Scholar] [CrossRef]
- Kipfmueller, K.F.; Baker, W.L. Fires and dwarf mistletoe in a Rocky Mountain lodgepole pine ecosystem. For. Ecol. Manag. 1998, 108, 77–84. [Google Scholar] [CrossRef]
- Manion, P. Tree Diseases Concepts, 2nd ed.; Prentice-Hall: Englewood Cliffs, NJ, USA, 1991. [Google Scholar]
- Knight, D.H. Parasites, lightning, and the vegetation mosaic in wilderness landscapes. In Landscape Heterogeneity and Disturbance; Springer: Berlin/Heidelberg, Germany, 1987; pp. 59–83. [Google Scholar]
- Conklin, D.A.; Armstrong, W.A. Effects of Three Prescribed Fires on Dwarf Mistletoe Infection in Southwestern Ponderosa Pine; US Department of Agriculture, Forest Service, Southwestern Region: Berkeley CA, USA, 2001. [Google Scholar]
- Haikerwal, A.; Reisen, F.; Sim, M.R.; Abramson, M.J.; Meyer, C.P.; Johnston, F.H.; Dennekamp, M. Impact of smoke from prescribed burning: Is it a public health concern? J. Air Waste Manag. Assoc. 2015, 65, 592–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afrin, S.; Garcia-Menendez, F. Potential impacts of prescribed fire smoke on public health and socially vulnerable populations in a Southeastern, U.S. state. Sci. Total Environ. 2021, 794, 148712. [Google Scholar] [CrossRef] [PubMed]
- Cirocco, R.M.; Facelli, J.M.; Watling, J.R. High water availability increases the negative impact of a native hemiparasite on its non-native host. J. Exp. Bot. 2015, 67, 1567–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griebel, A.; Metzen, D.; Pendall, E.; Nolan, R.H.; Clarke, H.; Renchon, A.A.; Boer, M.M. Recovery from Severe Mistletoe Infection After Heat- and Drought-Induced Mistletoe Death. Ecosystems 2022, 25, 1–16. [Google Scholar] [CrossRef]
- Griebel, A.; Peters, J.M.R.; Metzen, D.; Maier, C.; Barton, C.V.M.; Speckman, H.N.; Boer, M.M.; Nolan, R.H.; Choat, B.; Pendall, E. Tapping into the physiological responses to mistletoe infection during heat and drought stress. Tree Physiol. 2021, 42, 523–536. [Google Scholar] [CrossRef]
- Sidahmed, O.A. Incidence of mistletoe (Loranthus spp.) on citrus and guava trees in the central region of the Sudan. In Proceedings of the VIII African Symposium on Horticultural Crops 143, Wed Medani, Sudan, 20–24 March 1983; pp. 417–420. [Google Scholar]
- Knutson, D.M. The influence of urea fertilization on growth of mistle-toed pine seedlings. In Proceedings of the Second International Congress of plant pathology, Minneapolis, MN, USA, 5–12 September 1973; pp. 5–12. [Google Scholar]
- Kulkarni, H.D.; Srimathi, K. Plant pests of sandal (Santalum album L.). My For. 1988, 24, 29–38. [Google Scholar]
- Minko, G.; Fagg, P.C. Control of some mistletoe species on eucalypts by trunk injection with herbicides. Aust. For. 1989, 52, 94–102. [Google Scholar] [CrossRef]
- Brown, A.; Greenham, C. Further investigations in the control of mistletoe by trunk injections. Aust. J. Exp. Agric. 1965, 5, 305–309. [Google Scholar] [CrossRef]
- Brown, A. Mistletoe control on a large scale. J. Aust. Inst. Agric. Sci. 1959, 25, 282–286. [Google Scholar]
- Reid, N.; Fittler, J.; Kar, A.; Storey, A.; Cook, T. Effect of Spray Applications of Several Herbicides on the Mortality of Box Mistletoe (Amyema miquelii) and Host Saplings (Eucalyptus blakelyi and E. melliodora). In Ecosystem Management; University of New England: Armidale, Australia, 2008. [Google Scholar]
- Watson, W.T.; Martinez-Trinidad, T. Strategies and treatments for leafy mistletoe (Phoradendron tomentosum [DC.] Engelm ex. Gray) suppression on cedar elm (Ulmus crassifolia Nutt.). Arboric. Urban For. 2006, 32, 265. [Google Scholar] [CrossRef]
- Quick, C.R. Experimental Herbicidal Control of Dwarfmistletoe on Some California Conifers; U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1964; Volume 47, 9p. [Google Scholar]
- Livingston, W.; Brenner, M. Ethephon stimulates abscission of eastern dwarf mistletoe aerial shoots on black spruce. Plant Dis. 1983, 67, 909–910. [Google Scholar] [CrossRef]
- Livingston, W.H.; Blanchette, R.A.; Brenner, M.L.; Zuzek, K.J. Effective use of ethylene-releasing agents to prevent spread of eastern dwarf mistletoe on black spruce. Can. J. For. Res. 1985, 15, 872–876. [Google Scholar] [CrossRef]
- Livingston, W.H.; Brenner, M.L.; Blanchette, R.A. Altered concentrations of abscisic acid, indole-3-acetic acid, and zeatin riboside associated eastern dwarf mistletoe infections on black spruce. In Proceedings of the Biology of Dwarf Mistletoes, Symposium Proceedings, Fort Collins, CO, USA, 8 August 1984; Gen. Tech. Rep. RM-111. Hawksworth, F.G., Scharpf, R.F., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Berkeley, CA, USA, 1984; pp. 53–60. [Google Scholar]
- Adams, D.H.; Frankel, S.J.; Lichter, J.M. Considerations when using ethephon for supressing dwarf and leafy mistletoe infestations in ornamental landscapes. J. Arboric. 1993, 19, 351–357. [Google Scholar]
- Winder, R.S.; Shamoun, S.F. Forest pathogens: Friend or foe to biodiversity? Can. J. Plant Pathol. 2006, 28, S221–S227. [Google Scholar] [CrossRef]
- Robbins, K.; Johnson, D.W.; Hawksworth, F.G.; Nicholls, T.H. Aerial application of ethephon is ineffective in controlling lodgepole pine dwarf mistletoe. West. J. Appl. For. 1989, 4, 27–28. [Google Scholar] [CrossRef]
- Baker, F.; Knowles, K.; Meyer, T.; French, D. Aerial applications of ethylene-releasing chemicals fail to promote abscission of dwarf mistletoe aerial shoots on Jack pine. For. Chron. 1989, 65, 194–195. [Google Scholar] [CrossRef] [Green Version]
- Weber, H.C.; Forstreuter, W. Parasitismus von Blutenpflanzen; Wissenschaftliche Buchgesellschaft: Darmstadt, Germany, 1993. [Google Scholar]
- Parks, C.A.; Hoffman, J.T. Control of Western Dwarf Mistletoe with the Plant-Growth Regulator Ethephon; US Department of Agriculture, Forest Service, Pacific Northwest Research Station: Berkeley, CA, USA, 1991; Volume 506. [Google Scholar]
- Muir, J.; Geils, B. Management Strategies for Dwarf Mistletoe: Silviculture; Forest Service: Washington, DC, USA; USDA: Washington, DC, USA, 2002; pp. 83–94. [Google Scholar]
- Robinson, D.C.; Geils, B.W.; Muir, J.A. Spatial statistical model for the spread of dwarf mistletoe within and between stands. In Proceedings of the Second Forest Vegetation Simulator Conference, Fort Collins, CO, USA, 12–14 February 2002; Crookston, N.L., Havis, R.N., Eds.; pp. 178–185. [Google Scholar]
- Van Halder, I.; Castagneyrol, B.; Ordóñez, C.; Bravo, F.; del Río, M.; Perrot, L.; Jactel, H. Tree diversity reduces pine infestation by mistletoe. For. Ecol. Manag. 2019, 449, 117470. [Google Scholar] [CrossRef]
- Hanover, J.W. Tree improvement for disease resistance in western United States and Canada. In Breeding Pest-Resistant Trees; Elsevier: Amsterdam, The Netherlands, 1966; pp. 53–56. [Google Scholar]
- Geza, F.; Laszlo, J.; Ildiko, V.; Szilvia, P. Parasitic fungi of European mistletoe (Viscum album L.). Növényvédelem 2009, 45, 178–183. [Google Scholar]
- Scharpf, R.F. Host resistance to dwarf mistletoe. In Proceedings of the Biology of Dwarf Mistletoes, Symposium Proceedings, Fort Collins, CO, USA, 8 August 1984; Hawksworth, F.G., Scharpf, R.F., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Berkeley, CA, USA, 1984; pp. 70–76. [Google Scholar]
- Wyckhuys, K.A.G.; Hughes, A.C.; Buamas, C.; Johnson, A.C.; Vasseur, L.; Reymondin, L.; Deguine, J.P.; Sheil, D. Biological control of an agricultural pest protects tropical forests. Commun. Biol. 2019, 2, 10. [Google Scholar] [CrossRef]
- Nzioki, H.S.; Oyosi, F.; Morris, C.E.; Kaya, E.; Pilgeram, A.L.; Baker, C.S.; Sands, D.C. Striga Biocontrol on a Toothpick: A Readily Deployable and Inexpensive Method for Smallholder Farmers. Front. Plant Sci. 2016, 7, 1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bale, J.; Van Lenteren, J.; Bigler, F. Biological control and sustainable food production. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 761–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charudattan, R. Biological control of weeds by means of plant pathogens: Significance for integrated weed management in modern agro-ecology. BioControl 2001, 46, 229–260. [Google Scholar] [CrossRef]
- Hawksworth, F.G.; Geils, B.W. Biotic Associates; Diane Publishing: Washington, DC, USA; Department of Agriculture, Forest Service: Washington, DC, USA, 1996. [Google Scholar]
- Hawksworth, F.G.; Wicker, E.F.; Scharpf, R.F. Fungal Parasites of Dwarf Mistletoes [Arceuthobium, Parasites of Conifers, Biological Control]; Department of Agriculture, Forest Service: Washington, DC, USA, 1977; p. 14. [Google Scholar]
- Kuijt, J. Distribution of Dwarf Mistletoes and Their Fungus Hyperparasites in Western Canada; Department of Northern Affairs and National Resources: Ottawa, ON, Canada, 1963; Volume 186, pp. 134–148. [Google Scholar]
- Stevens, R.E.; Hawksworth, F.G. Insect-dwarf mistletoe associations: An update. In Proceedings of the Symposium: Biology of Dwarf Mistletoes, Fort Collins, CO, USA, 8 August 1984; pp. 94–101. [Google Scholar]
- Stevens, R.E.; Hawksworth, F.G. Insects and Mites Associated with Dwarf Mistletoes; Research Papers; Rocky Mountain Forest and Range Experiment Station: Fort Collins, CA, USA, 1970; p. 12. [Google Scholar]
- Ridings, W. Biological control of stranglervine in citrus—A researcher’s view. Weed Sci. 1986, 34, 31–32. [Google Scholar] [CrossRef]
- Daniel, J.; Templeton, G.; Smith, R.; Fox, W. Biological Control of Northern Jointvetch in Rice by An Endemic Fungal Disease. Weed Sci. 1973, 21, 303–307. [Google Scholar] [CrossRef]
- Makowski, R.M.; Mortensen, K. The first mycoherbicide in Canada: Colletotrichum gloeosporioides f. sp. malvae for round-leaved mallow control. In Proceedings of the First International Weed Control Congress, Melbourne, Australia, 17–21 February 1992; pp. 298–300. [Google Scholar]
- De Jong, M.; Scheepens, P.; Zadoks, J. Risk analysis for biological control: A Dutch case study in biocontrol of Prunus serotina by the fungus Chondrostereum purpureum. Plant Dis. 1990, 74, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Wall, R. Biological control of red alder using stem treatments with the fungus Chondrostereum purpureum. Can. J. For. Res. 1994, 24, 1527–1530. [Google Scholar] [CrossRef]
- Shamoun, S.F. Application of biological control to vegetation management in forestry. In Proceedings of the X International Symposium on Biological Control of Weeds, Bozeman, MO, USA, 4–14 July 1999; pp. 87–96. [Google Scholar]
- Shamoun, S. Implementation of weed biocontrol in forest vegetation management for conifer production. In Handbook of Sustainable Weed Management; Singh, H.P., Batish, D.R., Kohli, R.K., Eds.; Howarth Press, Inc.: Binghamton, NY, USA, 2006; pp. 475–506. [Google Scholar]
- Morris, M.; Wood, A.; Den Breeyen, A. Development and registration of a fungal inoculant to prevent regrowth of cut wattle tree stumps in South Africa and brief overview of other bioherbicide projects currenlty in progress. In Proceedings of the Fourth International Bioherbicide Workshop Programme and Abstracts, Glasgow, UK, 6–7 August 1998; University of Strathclyde: Strathclyde, UK, 1998; p. 15. [Google Scholar]
- Beilharz, V. Fungal pathogens of mistletoes in Victoria. Vic. Nat. 1997, 114, 162–163. [Google Scholar]
- Shamoun, S.F.; Ramsfield, T.D.; Van Der Kamp, B.J. Biological control approach for management of dwarf mistletoes. N. Z. J. For. Sci. 2003, 33, 373–384. [Google Scholar]
- Deeks, S.J.; Shamoun, S.F.; Punja, Z.K. In vitro germination and development of western hemlock dwarf mistletoe. Plant Cell Tissue Organ Cult. 2001, 66, 97–105. [Google Scholar] [CrossRef]
- Funk, A.; Smith, R.; Baranyay, J. Canker of dwarf mistletoe swellings on western hemlock caused by Nectria funkeliana var. macrospora. Can. J. For. Res. 1973, 3, 71–74. [Google Scholar] [CrossRef]
- Rietman, L.M.; van der Kamp, B.J.; Shamoun, S.F. Assessment of Neonectria neomacrospora (anamorph Cylindrocarpon cylindroides) as an inundative biocontrol agent against hemlock dwarf mistletoe. Can. J. Plant Pathol. 2005, 27, 603–609. [Google Scholar] [CrossRef]
- Scharpf, R.F. Dwarf Mistletoe on Red Fir: Infection and Control in Understory Stands; US Department of Agriculture, Pacific Southwest Forest and Range: Berkeley, CA, USA, 1969; Volume 50. [Google Scholar]
- Scharpf, R.F.; Bynum, H. Cytospora Canker of True Firs; Forest Service, US Department of Agriculture: Berkeley, CA, USA, 1975; Volume 146. [Google Scholar]
- Mark, W.R.; Hawksworth, F.G.; Oshima, N. Resin disease: A new disease of lodgepole pine dwarf mistletoe. Can. J. For. Res. 1976, 6, 415–424. [Google Scholar] [CrossRef]
- Kotan, R.; Okutucu, A.; Ala Görmez, A.; Karagoz, K.; Dadasoglu, F.; Karaman, I.; Hasanekoglu, I.; Kordali, Ş. Parasitic bacteria and fungi on common mistletoe (Viscum album L.) and their potential application in biocontrol. J. Phytopathol. 2013, 161, 165–171. [Google Scholar] [CrossRef]
- Ulukapı, M.; Erdoğdu, M. Aureobasidium Harposporum: A Potential Biocontrol Agent of Viscum Album. In Proceedings of the WITAM-2016, Kershehill, Turkey, 28 September–2 October 2016; p. 103. [Google Scholar]
- Mandeel, Q.; Hasan, A.; Al-Nafea, H.; Abbas, H. Antibacterial activity of extracts from selected marine algae in Bahrain. Arab Gulf J. Sci. Res. 2010, 28, 147–162. [Google Scholar] [CrossRef]
- Galal, H.; Salem, W.; Nasr El-Deen, F. Biological control of some pathogenic fungi using marine algae. Res. J. Microbiol 2011, 6, 645–657. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.A.; Khan, A.A. Use of diatomaceous earth for the management of stored-product pests. Int. J. Pest Manag. 2014, 60, 100–113. [Google Scholar] [CrossRef]
- El-Ansary, M.S.M.; Hamouda, R.A. Biocontrol of root-knot nematode infected banana plants by some marine algae. Russ. J. Mar. Biol. 2014, 40, 140–146. [Google Scholar] [CrossRef]
- Nabaei, N.; Mehrvar, A.; Saber, M.; Bagheri, M. Efficacy of entomopathogenic fungi in combination with diatomaceous earth against Callosobruchus maculatus (Coleoptera: Bruchidae). Acta Entomol. Sin. 2012, 55, 1282–1288. [Google Scholar]
- Athanassiou, C.G.; Palyvos, N.E. Laboratory evaluation of two diatomaceous earth formulations against Blattisocius keegani Fox (Mesostigmata, Ascidae) and Cheyletus malaccensis oudemans (Prostigmata, Cheyletidae). Biol. Control 2006, 38, 350–355. [Google Scholar] [CrossRef]
- Solomon, J.; Newsome, L.; Filer, T. Discovery and observations of a stem-boring weevil (Myrmex sp.) a potentially useful biocontrol of mistletoe. J. Miss. Acad. Sci. 1984, 29, 7–11. [Google Scholar]
- Van Dyke, E.C. New Rhynchophora (Coleoptera) from western North America. Pan-Pac. Entomol. 1930, 6, 149–165. [Google Scholar]
- Sleeper, E.L. New genera and species of Curculionidae with a new species of Anthribidae (Coleoptera). New Curculionidae 1953, 53, 113–120. [Google Scholar]
- Vanin, S.A.; Guerra, T.J. A remarkable new species of flesh-fly mimicking weevil (Coleoptera: Curculionidae: Conoderinae) from Southeastern Brazil. Zootaxa 2012, 3413, 55–63. [Google Scholar] [CrossRef]
- Guerra, T.J. Evasive mimicry: Too beetle, or not too beetle? Ecology 2019, 100, e02773. [Google Scholar] [CrossRef] [PubMed]
- Baloch, G.; Mohyuddin, A. The phytophagous fauna of a mistletoe (Loranthus longiflorus Desr.: Loranthaceae) in West Pakistan. Weed Res. 1969, 9, 62–64. [Google Scholar] [CrossRef]
- Mushtaque, M.; Baloch, G. Possibilities of biological control of mistletoes, Loranthus spp., using oligophagous insects from Pakistan. Entomophaga 1979, 24, 73–81. [Google Scholar] [CrossRef]
- Jendek, E.; Poláková, J. Host Plants of World Agrilus (Coleoptera, Buprestidae), 1st ed.; Springer: Cham, Germany, 2014; Volume 706. [Google Scholar]
- Królik, R. Agrilus kutahyanus n. sp. from Turkey (Coleoptera: Buprestidae). Genus 2002, 13, 25–31. [Google Scholar]
- Lastuvka, Z. Morphology and biology of clearwing moths Synanthedon loranthi (Kr.) and Synanthedon cephiformis (O.) (Lepidoptera, Sesiidae). Acta Univ. Agric. Fac. Agron 1983, 31, 143–158. [Google Scholar]
- Senn, P.; Kowalczyk, J. Nowe stanowiska Ditula angustiorana (Haworth, 1811)(Lepidoptera: Tortricidae) oraz pierwsze stwierdzenia samców tego gatunku w Polsce. Wiadomości Entomol. 2018, 37, 125–126. [Google Scholar]
- Buszko, J.; Nowacki, J.; Entomologiczne, P.T. A Distributional Checklist of the Lepidoptera of Poland; Polish Entomological Society: Poznan, Poland, 2017. [Google Scholar]
- Reichholf, J.H. Aspekte der Biologie und Fragen zur Verbreitung des Mistelwicklers Celypha woodiana Barrett, 1882, in Bayern. Atalanta 2008, 39, 379–383. [Google Scholar]
- Briggs, J. Mistletoe (Viscum album): A brief review of its local status with recent observations on its insects associations and conservation problems. In Proceedings of the Cotteswold Naturalists’ Field Club, Cotteswold, UK, 12 September 2011; pp. 181–193. [Google Scholar]
- Lazaro-Gonzalez, A.; Hodar, J.A.; Zamora, R. Do the arthropod communities on a parasitic plant and its hosts differ? Eur. J. Entomol. 2017, 114, 215. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Fernández-Aparicio, M. First report of cottony-cushion scale (Icerya purchasi) on red berried mistletoe (Viscum cruciatum). Entomol. Res. 2009, 39, 95–96. [Google Scholar] [CrossRef]
- Chávez-Salcedo, L.F.; Queijeiro-Bolaños, M.E.; López-Gómez, V.; Cano-Santana, Z.; Mejía-Recamier, B.E.; Mojica-Guzmán, A. Contrasting arthropod communities associated with dwarf mistletoes Arceuthobium globosum and A. vaginatum and their host Pinus hartwegii. J. For. Res. 2018, 29, 1351–1364. [Google Scholar] [CrossRef]
- Tong, J.; Ren, W. Preliminary studies on the disease cycle of Arceuthobium chinense. J. Yunnan For. Coll. 1980, 1, 19–25. [Google Scholar]
- Krasylenko, Y.; Těšitel, J.; Ceccantini, G.; Oliveira-da-Silva, M.; Dvořák, V.; Steele, D.; Sosnovsky, Y.; Piwowarczyk, R.; Watson, D.M.; Teixeira-Costa, L. Parasites on parasites: Hyper-, epi-, and autoparasitism among flowering plants. Am. J. Bot. 2021, 108, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Calvin, C.L.; Wilson, C.A. Epiparasitism in Phoradendron durangense and P. falcatum (Viscaceae). Aliso A J. Syst. Evol. Bot. 2009, 27, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Glatzel, G.; Balasubramaniam, S. Mineral nutrition of mistletoes: General concepts. In Proceedings of the 4th International Symposium on Parasitic Flowering Plants, Marburg, Germany, 10 August 1987; pp. 263–276. [Google Scholar]
- Wiens, D.; Calvin, C. Epiparasitism in mistletoes. Gold. Bough (R. Bot. Gard. Kew) 1987, 9, 3–5. [Google Scholar]
- Downey, P.O. An inventory of host species for each aerial mistletoe species (Loranthaceae and Viscaceae) in Australia. Cunninghamia 1998, 5, 685–720. [Google Scholar]
- Moss, J.S.L. Mistletoe/hostplant associations of the Barakula, Columboola and Gurulmundi region, south-east Queensland. Qld. Nat. 1998, 36, 73–87. [Google Scholar]
- Eastwood, K. A Guide to Australia’s Possums and Gliders. Available online: https://www.australiangeographic.com.au/topics/wildlife/2019/07/a-guide-to-all-27-species-of-australias-possums-and-gliders/ (accessed on 22 October 2022).
- Sessions, L.A.; Rance, C.; Grant, A.; Kelly, D. Possum (Trichosurus vulpecula) control benefits native beech mistletoes (Loranthaceae). N. Z. J. Ecol. 2001, 25, 27–33. [Google Scholar]
- Reid, N. Control of mistletoes by possums and fire: A review of the evidence. Vic. Nat. 1997, 114, 149–158. [Google Scholar]
- Kar, A.K. Methods to Manage Mistletoe—A Landholder’s Guide. 2008. Available online: https://www.snelandcare.org.au/linkedfiles/MethodsManageMistletoe.pdf (accessed on 8 October 2022).
- Room, P. The constitution and natural history of the fauna of the mistletoe Tapinanthus bangwensis (Engl. & K. Krause) growing on cocoa in Ghana. J. Anim. Ecol. 1972, 41, 519–535. [Google Scholar]
- Patrick, B.H.; Dugdale, J.S. Mistletoe moths-New Zealand’s loranthaceous mistletoes. In Proceedings of the A Workshop Hosted by Threatened Species Unit, Department of Conservation, Cass, Wellington, New Zealand, 17–20 July 1995; p. 125. [Google Scholar]
- Wicker, E.F.; Shaw, C.G. Fungal parasites of dwarf mistletoes. Mycologia 1968, 60, 372–383. [Google Scholar] [CrossRef]
- Nguyen, J.; Fernandez, V.; Pontrelli, S.; Sauer, U.; Ackermann, M.; Stocker, R. A distinct growth physiology enhances bacterial growth under rapid nutrient fluctuations. Nat. Commun. 2021, 12, 3662. [Google Scholar] [CrossRef] [PubMed]
- Burman, E.; Bengtsson-Palme, J. Microbial Community Interactions Are Sensitive to Small Changes in Temperature. Front. Microbiol. 2021, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hardy, D.E. Studies of fruitflies associated with mistletoe in Australia and Pakistan with notes and decriptions on genera related to Perilampsis Bezzi (Diptera: Tephritidae). Beiträge Zur Entomol. Contrib. Entomol. 1967, 17, 127–149. [Google Scholar]
- Garbelotto, M.; Lowell, N.; Chen, I.Y.; Osmundson, T.W. Evidence for inhibition of a fungal biocontrol agent by a plant microbiome. J. Plant Pathol. 2019, 101, 457–466. [Google Scholar] [CrossRef]
- Perry, E. Broadleaf Mistletoe in Landscape Trees; University of California: Berkeley, CA, USA, 1995; p. 14. [Google Scholar]
- Rubiales, D. Parasitic plants, wild relatives and the nature of resistance. New Phytol. 2003, 160, 459–461. [Google Scholar] [CrossRef]
- Aly, R.; Dubey, N.K. Weed Management for Parasitic Weeds. In Recent Advances in Weed Management; Springer: Berlin/Heidelberg, Germany, 2014; pp. 315–345. [Google Scholar]
- Hawksworth, F.G. Biological factors of dwarf mistletoe in relation to control. In Proceedings of the Symposium on Dwarf Mistletoes Control through Forest Management, Berkeley, CA, USA, 11–13 April 1978; Gen. Tech. Rep. PSW-31. pp. 5–15. [Google Scholar]
- Roth, L.F. Genetic control of dwarf mistletoe. In Proceedings of the Symposium on Dwarf Mistletoes Control through Forest Management, Berkeley, CA, USA, 11–13 April 1978; pp. 69–72. [Google Scholar]
- Scharpf, R.F.; Roth, L.F. Resistance of Ponderosa Pine to Western Dwarf Mistletoe in Central Oregon; Res. Pap. PSW-RP-208; US Department of Agriculture, Pacific Southwest Forest and Range: Albany, CA, USA, 1992; Volume 208, 9p. [Google Scholar]
- Roth, L.F. Resistance of ponderosa pine to dwarf mistletoe. Silvae Genet. 1974, 23, 116–120. [Google Scholar]
- Arruda, R.; Carvalho, L.N.; Del-Claro, K. Host specificity of a Brazilian mistletoe, Struthanthus aff. polyanthus (Loranthaceae), in Cerrado Tropical Savanna. Flora-Morphol. Distrib. Funct. Ecol. Plants 2006, 201, 127–134. [Google Scholar] [CrossRef]
- Atsatt, P. Host-parasite interactions in higher plants. In Physiological Plant Ecology III; Springer: Berlin/Heidelberg, Germany, 1983; Volume 12/C, pp. 519–535. [Google Scholar]
- Dean, W.; Midgley, J.; Stock, W. The distribution of mistletoes in South Africa: Patterns of species richness and host choice. J. Biogeogr. 1994, 21, 503–510. [Google Scholar] [CrossRef]
- Downey, P.O.; Gill, A.M.; Banks, J.C. The Influence of Host Attributes on Mistletoe Colonization: An Example from Mulligan’s Flat Nature Reserve, A.C.T. Vic. Nat. 1997, 114, 105–111. [Google Scholar]
- Ghosh, S.K.; Balasundaram, M.; Mohamed, A.M. Studies in Host Parasite Relationship of Phanerogamic Parasites on Teak and Their Possible Control; Kerela Forest Reseach Institute: Kerala, India, 1984; p. 21. [Google Scholar]
- Halbritter, D.A.; Willett, D.S.; Gordon, J.M.; Stelinski, L.L.; Daniels, J.C. Behavioral Evidence for Host Transitions in Plant, Plant Parasite, and Insect Interactions. Environ. Entomol. 2018, 47, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.J.; Fuentes, E.R.; Cortes, I.; Liberona, F.; Costa, V. Tristerix tetrandrus (Loranthaceae) and its host-plants in the Chilean matorral: Patterns and mechanisms. Oecologia 1986, 69, 202–206. [Google Scholar] [CrossRef]
- Norton, D.A.; Carpenter, M.A. Mistletoes as parasites: Host specificity and speciation. Trends Ecol. Evol. 1998, 13, 101–105. [Google Scholar] [CrossRef]
- Okubamichael, D.Y.; Griffiths, M.E.; Ward, D. Host specificity in parasitic plants—Perspectives from mistletoes. AoB Plants 2016, 8, plw069. [Google Scholar] [CrossRef]
- Rödl, T.; Ward, D. Host recognition in a desert mistletoe: Early stages of development are influenced by substrate and host origin. Funct. Ecol. 2002, 16, 128–134. [Google Scholar] [CrossRef]
- Tainter, F.; French, D. The role of wound periderm in the resistance of eastern larch and jack pine to dwarf mistletoe. Can. J. Bot. 1971, 49, 501–504. [Google Scholar] [CrossRef]
- Smith, R.; Wass, E.; Meagher, M. Evidence of resistance to hemlock dwarf mistletoe (Arceuthobium tsugense) in western hemlock (Tsuga heterophylla) clones. Eur. J. For. Pathol. 1993, 23, 163–170. [Google Scholar] [CrossRef]
- Roeser, J., Jr. The importance of seed source and the possibilities of forest tree breeding. J. For. 1926, 24, 38–51. [Google Scholar] [CrossRef]
- Bates, C.G. Better seeds, better trees. J. For. 1927, 25, 130–144. [Google Scholar] [CrossRef]
- Hawksworth, F.G.; Edminster, C.B. Carlos Bates’ Dwarf Mistletoe Resistant Ponderosa Pines: A Postscript after Half a Century; USDA Forest Service, Rocky Mountain Forest and Range Experiment Station: Berkeley, CA, USA, 1981; Volume 412, p. 4. [Google Scholar]
- Roth, L.F. Dwarf mistletoe damage to small ponderosa pines. For. Sci. 1971, 17, 373–380. [Google Scholar] [CrossRef]
- Roth, L.F. Juvenile Susceptibility of Ponderosa Pine. Phytopathology 1974, 64, 689–692. [Google Scholar] [CrossRef]
- Harrington, T.; Wingfield, M. Diseases and the ecology of indigenous and exotic pines. In Ecology and Biogeography of Pinus; Richardson, D., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 1–33. [Google Scholar]
- Okubamichael, D.Y. Host Specificity of the Hemiparasitic Mistletoe, Agelanthus natalitius. Master’s Thesis, University of KwaZulu-Natal, Pietermaritzburg, South Africa, 2009. [Google Scholar]
- Skrypnik, L.; Maslennikov, P.; Feduraev, P.; Pungin, A.; Belov, N. Ecological and Landscape Factors Affecting the Spread of European Mistletoe (Viscum album L.) in Urban Areas (A Case Study of the Kaliningrad City, Russia). Plants 2020, 9, 394. [Google Scholar] [CrossRef] [Green Version]
- Zaroug, M.S.; Zahran, E.B.; Abbasher, A.A. Distribution And Host Range of Mistletoe (Tapinanthus globiferus) (A. Rich.) Van Tieghan) Along the Blue Nile Banks in Central Sudan. Int. J. Sci. Technol. Res. 2014, 3, 1–5. [Google Scholar]
- Ladley, J.J.; Kelly, D.; Robertson, A.W. Explosive flowering, nectar production, breeding systems, and pollinators of New Zealand mistletoes (Loranthaceae). N. Z. J. Bot. 1997, 35, 345–360. [Google Scholar] [CrossRef] [Green Version]
- Lavorel, S.; Smith, M.S.; Reid, N. Spread of mistletoes (Amyema preissii) in fragmented Australian woodlands: A simulation study. Landsc. Ecol. 1999, 14, 147–160. [Google Scholar] [CrossRef]
- Yoder, J.I. Parasitic plant responses to host plant signals: A model for subterranean plant–plant interactions. Curr. Opin. Plant Biol. 1999, 2, 65–70. [Google Scholar] [CrossRef]
- Tomilov, A.; Tomilova, N.; Shin, D.H.; Jamison, D.; Torres, M.; Reagan, R.; McGray, H.; Horning, T.; Truong, R.; Nava, A. Chemical signalling between plants. In Chemical Ecology: From Gene to Ecosystem; Springer: Dordrecht, The Netherlands, 2006; Volume 16, pp. 55–69. [Google Scholar]
- Yan, Z. Resistance to haustorial development of two mistletoes, Amyema preissii (Miq.) Tieghem and Lysiana exocarpi (Behr.) Tieghem ssp. exocarpi (Loranthaceae), on host and nonhost species. Int. J. Plant Sci. 1993, 154, 386–394. [Google Scholar] [CrossRef]
- Lech, P.; Żółciak, A.; Hildebrand, R. Occurrence of European Mistletoe (Viscum album L.) on Forest Trees in Poland and Its Dynamics of Spread in the Period 2008–2018. Forests 2020, 11, 83. [Google Scholar] [CrossRef]
- Nickrent, D.; Musselman, L. Introduction to parasitic flowering plants. Plant Health Instr. 2004. [Google Scholar] [CrossRef]
- Overton, J.M. Dispersal and infection in mistletoe metapopulations. J. Ecol. 1994, 82, 711–723. [Google Scholar] [CrossRef]
- Amico, G.; Aizen, M.A. Mistletoe seed dispersal by a marsupial. Nature 2000, 408, 929–930. [Google Scholar] [CrossRef]
- Lira-Noriega, A.; Toro-Núñez, O.; Oaks, J.R.; Mort, M.E. The roles of history and ecology in chloroplast phylogeographic patterns of the bird-dispersed plant parasite Phoradendron californicum (Viscaceae) in the Sonoran Desert. Am. J. Bot. 2015, 102, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Crespo, M.; Ornelas, J.; Martén-Rodríguez, S.; González-Rodríguez, A.; Lara, C. Reproductive biology and nectar production of the Mexican endemic P. sittacanthus auriculatus (Loranthaceae), a hummingbird-pollinated mistletoe. Plant Biol. 2016, 18, 73–83. [Google Scholar] [CrossRef]
- Burgess, V.J.; Kelly, D.; Robertson, A.W.; Ladley, J.J. Positive effects of forest edges on plant reproduction: Literature review and a case study of bee visitation to flowers of Peraxilla tetrapetala (Loranthaceae). N. Z. J. Ecol. 2006, 30, 179–190. [Google Scholar]
- Kelly, D.; Ladley, J.J.; Robertson, A.W.; Crowfoot, L. Flower predation by Zelleria maculata (Lepidoptera) on Peraxilla mistletoes: Effects of latitude and fragmentation, and impact on fruit set. N. Z. J. Ecol. 2008, 32, 186–196. [Google Scholar]
- MacRaild, L.M.; Radford, J.Q.; Bennett, A.F. Non-linear effects of landscape properties on mistletoe parasitism in fragmented agricultural landscapes. Landsc. Ecol. 2010, 25, 395–406. [Google Scholar] [CrossRef]
- Bowen, M.E.; McAlpine, C.A.; House, A.P.; Smith, G.C. Agricultural landscape modification increases the abundance of an important food resource: Mistletoes, birds and brigalow. Biol. Conserv. 2009, 142, 122–133. [Google Scholar] [CrossRef]
- Reid, N.; Stafford Smith, M.; Yan, Z. Ecology and population biology of mistletoes. In Forest Canopies; Lowman, M.D., Nadkarn, N.M., Eds.; Elsevier: Amsterdam, The Netherlands, 1995; pp. 285–310. [Google Scholar]
- Norton, D.A.; Ladley, J.J. Establishment and early growth of Alepis flavida in relation to Nothofagus solandri branch size. N. Z. J. Bot. 1998, 36, 213–217. [Google Scholar] [CrossRef]
- Aukema, J.E.; Del Rio, C.M. Variation in mistletoe seed deposition: Effects of intra-and interspecific host characteristics. Ecography 2002, 25, 139–144. [Google Scholar] [CrossRef]
- Baena-Díaz, F.; Ramírez-Barahona, S.; Ornelas, J.F. Hybridization and differential introgression associated with environmental shifts in a mistletoe species complex. Sci. Rep. 2018, 8, 5591. [Google Scholar] [CrossRef] [PubMed]
- Azpeitia, F.; Lara, C. Reproductive biology and pollination of the parasitic plant Psittacanthus calyculatus (Loranthaceae) in central México1. J. Torrey Bot. Soc. 2006, 133, 429–438. [Google Scholar] [CrossRef]
- Ramírez, M.M.; Ornelas, J.F. Polinización y producción de néctar de Psittacanthus schiedeanus (Loranthaceae) en el centro de Veracruz, México. Boletín De La Soc. Botánica De México 2010, 87, 61–67. [Google Scholar]
- Guerra, T.; Galetto, L.; Silva, W. Nectar secretion dynamic links pollinator behavior to consequences for plant reproductive success in the ornithophilous mistletoe Psittacanthus robustus. Plant Biol. 2014, 16, 956–966. [Google Scholar] [CrossRef]
- Belchior, M.M.; Camarota, F.; Antiqueira, P.A.P.; Neves, F.S. A neotropical mistletoe influences herbivory of its host plant by driving changes in the associated insect community. Sci. Nat. 2022, 109, 27. [Google Scholar] [CrossRef]
- Ollerton, J.; Stott, A.; Allnutt, E.; Shove, S.; Taylor, C.; Lamborn, E. Pollination niche overlap between a parasitic plant and its host. Oecologia 2007, 151, 473–485. [Google Scholar] [CrossRef]
- Kuijt, J. Monograph of Psittacanthus (Loranthaceae). Syst. Bot. Monogr. 2009, 86, 1–362. [Google Scholar]
- Amico, G.C.; Vidal-Russell, R.; Garcia, M.A.; Nickrent, D.L. Evolutionary history of the South American mistletoe Tripodanthus (Loranthaceae) using nuclear and plastid markers. Syst. Bot. 2012, 37, 218–225. [Google Scholar] [CrossRef]
- Ornelas, J.F.; Gándara, E.; Vásquez-Aguilar, A.A.; Ramírez-Barahona, S.; Ortiz-Rodriguez, A.E.; González, C.; Saules, M.T.M.; Ruiz-Sanchez, E. A mistletoe tale: Postglacial invasion of Psittacanthus schiedeanus (Loranthaceae) to Mesoamerican cloud forests revealed by molecular data and species distribution modeling. BMC Evol. Biol. 2016, 16, 78. [Google Scholar] [CrossRef] [Green Version]
- Reid, N. Dispersal of misteltoes by honeyeaters and flowerpeckers: Components of seed dispersal quality. Ecology 1989, 70, 137–145. [Google Scholar] [CrossRef]
- Godschalk, S. Feeding behaviour of avian dispersers of mistletoe fruit in the Loskop Dam Nature Reserve, South Africa. Afr. Zool. 1985, 20, 136–146. [Google Scholar] [CrossRef]
- Van Ommeren, R.J.; Whitham, T.G. Changes in interactions between juniper and mistletoe mediated by shared avian frugivores: Parasitism to potential mutualism. Oecologia 2002, 130, 281–288. [Google Scholar] [CrossRef] [PubMed]
- García, D.; Rodríguez-Cabal, M.A.; Amico, G.C. Seed dispersal by a frugivorous marsupial shapes the spatial scale of a mistletoe population. J. Ecol. 2009, 97, 217–229. [Google Scholar] [CrossRef]
- Rodriguez-Cabal, M.A.; Branch, L.C. Influence of habitat factors on the distribution and abundance of a marsupial seed disperser. J. Mammal. 2011, 92, 1245–1252. [Google Scholar] [CrossRef] [Green Version]
- Morales, J.M.; Rivarola, M.D.; Amico, G.; Carlo, T.A. Neighborhood effects on seed dispersal by frugivores: Testing theory with a mistletoe–marsupial system in Patagonia. Ecology 2012, 93, 741–748. [Google Scholar] [CrossRef] [Green Version]
- Arriaga, L.; Franco, M.; Sarukhan, J. Identification of natural groups of trees in uneven-aged forests using multivariate methods. J. Ecol. 1988, 76, 1092–1100. [Google Scholar] [CrossRef]
- Donohue, K. The spatial demography of mistletoe parasitism on a Yemeni Acacia. Int. J. Plant Sci. 1995, 156, 816–823. [Google Scholar] [CrossRef]
- Gairola, S.; Bhatt, A.; Govender, Y.; Baijnath, H.; Procheş, Ş.; Ramdhani, S. Incidence and intensity of tree infestation by the mistletoe Erianthemum dregei (Eckl. & Zeyh.) V. Tieghem in Durban, South Africa. Urban For. Urban Green. 2013, 12, 315–322. [Google Scholar] [CrossRef]
- Teixeira-Costa, L.; Coelho, F.M.; Ceccantini, G.C.T. Comparative phenology of mistletoes shows effect of different host species and temporal niche partitioning. Botany 2017, 95, 271–282. [Google Scholar] [CrossRef]
- Yule, K.M.; Bronstein, J.L. Reproductive ecology of a parasitic plant differs by host species: Vector interactions and the maintenance of host races. Oecologia 2018, 186, 471–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuadra-Valdés, J.; Vizentin-Bugoni, J.; Fontúrbel, F.E. An exotic magnet plant alters pollinator abundance and behavior: A field test with a native mistletoe. Biol. Invasions 2021, 23, 2515–2525. [Google Scholar] [CrossRef]
- Watson, D.M. Parasitic plants as facilitators: More Dryad than Dracula? J. Ecol. 2009, 97, 1151–1159. [Google Scholar] [CrossRef]
- Watson, D.M. Disproportionate declines in ground-foraging insectivorous birds after mistletoe removal. PLoS ONE 2015, 10, e0142992. [Google Scholar] [CrossRef]
- Hódar, J.A.; Lázaro-González, A.; Zamora, R. Beneath the mistletoe: Parasitized trees host a more diverse herbaceous vegetation and are more visited by rabbits. Ann. For. Sci. 2018, 75, 77. [Google Scholar] [CrossRef] [Green Version]
- Mellado, A.; Hobby, A.; Lázaro-González, A.; Watson, D.M. Hemiparasites drive heterogeneity in litter arthropods: Implications for woodland insectivorous birds. Austral Ecol. 2019, 44, 777–785. [Google Scholar] [CrossRef]
- Lázaro-González, A.; Hódar, J.A.; Zamora, R. Mistletoe generates non-trophic and trait-mediated indirect interactions through a shared host of herbivore consumers. Ecosphere 2019, 10, e02564. [Google Scholar] [CrossRef]
- Ferrenberg, S. Dwarf Mistletoe Infection Interacts with Tree Growth Rate to Produce Opposing Direct and Indirect Effects on Resin Duct Defenses in Lodgepole Pine. Forests 2020, 11, 222. [Google Scholar] [CrossRef] [Green Version]
- Zamora, R.; Mellado, A. Identifying the abiotic and biotic drivers behind the elevational distribution shift of a parasitic plant. Plant Biol. 2019, 21, 307–317. [Google Scholar] [CrossRef]
- Lawson, D.A.; Rands, S.A. The effects of rainfall on plant–pollinator interactions. Arthropod-Plant Interact. 2019, 13, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Cubero, J.; Hernández, L. Breeding faba bean (Vicia faba L.) for resistance to Orobanche crenata Forsk. Options Méditerranéennes 1991, 10, 51–57. [Google Scholar]
- Ringnes, D.; Stover, P.; Scharpf, R. Dwarf mistletoe resistance in ponderosa pine: Selection and testing protocols. In Proceedings of the Congresos y Jornadas-Junta de Andalucia (Espana), Cordoba, Spain, 16–18 April 1996; pp. 690–696. [Google Scholar]
- Haussmann, B.; Hess, D.; Reddy, B.; Mukuru, S.; Seetharama, N.; Kayentao, M.; Welz, H.; Geiger, H. QTL for Striga resistance in sorghum populations derived from IS 9830 and N 13. In Proceedings of the Breeding for Striga Resistance in Cereals: Proceedings of a Workshop, IITA, Ibadan, Nigeria, 18–20 August 1999; pp. 18–20. [Google Scholar]
- Mohamed, A.; Rich, P.; Housley, T.; Ejeta, G. In vitro techniques for studying mechanisms of Striga resistance in sorghum. In Proceedings of the 7th International Parasitic Weed Symposium, Nantes, France, 3–8 June 2001; pp. 96–100. [Google Scholar]
- Omanya, G.O.; Haussmann, B.I.G.; Hess, D.E.; Reddy, B.V.S.; Kayentao, M.; Welz, H.G.; Geiger, H.H. Utility of indirect and direct selection traits for improving Striga resistance in two sorghum recombinant inbred populations. Field Crops Res. 2004, 89, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, B.; Guo, Q.; Song, L.; Chen, L.; Wang, C. Construction of a haustorium development associated SSH library in Thesium chinense and analysis of specific ESTs included by Imperata cylindrica. Biochem. Syst. Ecol. 2016, 64, 46–52. [Google Scholar] [CrossRef]
- Ranjan, A.; Ichihashi, Y.; Farhi, M.; Zumstein, K.; Townsley, B.; David-Schwartz, R.; Sinha, N.R. De novo assembly and characterization of the transcriptome of the parasitic weed dodder identifies genes associated with plant parasitism. Plant Physiol. 2014, 166, 1186–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichihashi, Y.; Kusano, M.; Kobayashi, M.; Suetsugu, K.; Yoshida, S.; Wakatake, T.; Kumaishi, K.; Shibata, A.; Saito, K.; Shirasu, K.J.P.; et al. Transcriptomic and metabolomic reprogramming from roots to haustoria in the parasitic plant, Thesium chinense. Plant Cell Physiol. 2018, 59, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Zhou, W.; Li, T.; Tian, C.J.B.g. De novo assembly and transcriptome characterization of spruce dwarf mistletoe Arceuthobium sichuanense uncovers gene expression profiling associated with plant development. BMC Genom. 2016, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Abdul Wahid, H. Identification of Dwarf Mistletoe Resistant Genes in Ziarat Junipers (Juniperus excelsa M. Bieb). Ph.D. Thesis, University of Baluchistan, Quetta, Pakistan, 2019. [Google Scholar]
- Abdul Wahid, H.; Barozai, M.Y.K.; Din, M. Identification and characterization of dwarf mistletoe responding genes in Ziarat juniper tree (Juniperus excelsa M. Bieb) through suppression subtractive hybridization and deep sequencing. Trees 2019, 33, 1027–1039. [Google Scholar] [CrossRef]
- Kuang, J.; Wang, Y.; Mao, K.; Milne, R.; Wang, M.; Miao, N. Transcriptome Profiling of a Common Mistletoe Species Parasitizing Four Typical Host Species in Urban Southwest China. Genes 2022, 13, 1173. [Google Scholar] [CrossRef]
- Dan, M.W.; Filiz, G. Plant Genome Editing and its Applications in Cereals. In Genetic Engineering; Farrukh, J., Ed.; IntechOpen: Rijeka, Croatia, 2016; p. 4. [Google Scholar]
- Mandal, K.; Boro, P.; Chattopadhyay, S. Micro-RNA based gene regulation: A potential way for crop improvements. Plant Gene 2021, 27, 100312. [Google Scholar] [CrossRef]
- Yu, H.; Li, J. Breeding future crops to feed the world through de novo domestication. Nat. Commun. 2022, 13, 1171. [Google Scholar] [CrossRef] [PubMed]
- Van Esse, H.P.; Reuber, T.L.; van der Does, D. Genetic modification to improve disease resistance in crops. New Phytol. 2020, 225, 70–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.S.-e.-A.; Mahas, A.; Vanderschuren, H.; Mahfouz, M.M. Engineering crops of the future: CRISPR approaches to develop climate-resilient and disease-resistant plants. Genome Biol. 2020, 21, 289. [Google Scholar] [CrossRef] [PubMed]
- Clarke, C.R.; Timko, M.P.; Yoder, J.I.; Axtell, M.J.; Westwood, J.H. Molecular Dialog Between Parasitic Plants and Their Hosts. Annu. Rev. Phytopathol. 2019, 57, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Westwood, J.H.; de Pamphilis, C.W.; Das, M.; Fernández-Aparicio, M.; Honaas, L.A.; Timko, M.P.; Wafula, E.K.; Wickett, N.J.; Yoder, J.I. The Parasitic Plant Genome Project: New Tools for Understanding the Biology of Orobanche and Striga. Weed Sci. 2017, 60, 295–306. [Google Scholar] [CrossRef]
- Neale, D.B.; Kremer, A. Forest tree genomics: Growing resources and applications. Nat. Rev. Genet. 2011, 12, 111. [Google Scholar] [CrossRef]
- Muleo, R.; Morgante, M.; Velasco, R.; Cavallini, A.; Perrotta, G.; Baldoni, L. Olive tree genomic. In Olive Germplasm–The Olive Cultivation, Table Olive and Olive Oil Industry in Italy, 1st ed.; Muzzalupo, I., Ed.; Intechopen: Rijeka, Croatia, 2012; pp. 133–148. [Google Scholar]
- Diningrat, D.; Widiyanto, S.; Pancoro, A.; Shim, D.; Panchangam, B.; Zembower, N.; Carlson, J.E. Transcriptome of teak (Tectona grandis, Lf) in vegetative to generative stages development. J. Plant Sci. 2015, 10, 1. [Google Scholar] [CrossRef]
- Schnell, R.J.; Priyadarshan, P. Genomics of Tree Crops; Springer Science & Business Media; Springer: New York, NY, USA, 2012. [Google Scholar]
- Gmitter, F.G.; Chen, C.; Machado, M.A.; De Souza, A.A.; Ollitrault, P.; Froehlicher, Y.; Shimizu, T. Citrus genomics. Tree Genet. Genomes 2012, 8, 611–626. [Google Scholar] [CrossRef] [Green Version]
- Arias, R.S.; Borrone, J.W.; Tondo, C.L.; Kuhn, D.N.; Irish, B.M.; Schnell, R.J. Genomics of tropical fruit tree crops. In Genomics of Tree Crops; Springer: Berlin/Heidelberg, Germany, 2012; pp. 209–239. [Google Scholar]
- Marler, M.; Pedersen, D.; Mitchell-Olds, T.; Callaway, R. A polymerase chain reaction method for detecting dwarf mistletoe infection in Douglas-fir and western larch. Can. J. For. Res. 1999, 29, 1317–1321. [Google Scholar] [CrossRef]
- Petri, C.; Burgos, L. Transformation of fruit trees. Useful breeding tool or continued future prospect? Transgenic Res. 2005, 14, 15–26. [Google Scholar] [CrossRef]
- Atsatt, P.R. On the evolution of leaf resemblance between mistletoes and their hosts. In Proceedings of the 2nd Symposium on Parasitic Weeds, Raleigh, NC, USA, 16–19 July 1979. [Google Scholar]
- Lev-Yadun, S. Does chemical aposematic (warning) signaling occur between host plants and their potential parasitic plants? Plant Signal. Behav. 2013, 8, e24907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, S.; Cui, S.; Ichihashi, Y.; Shirasu, K. The Haustorium, a Specialized Invasive Organ in Parasitic Plants. Annu. Rev. Plant Biol. 2016, 67, 643–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.; Lynn, D.G. The haustorium and the chemistry of host recognition in parasitic angiosperms. J. Chem. Ecol. 1986, 12, 561–579. [Google Scholar] [CrossRef]
- Sallé, G.L. Germination and Establishment of Viscum album. In The Biology of Mistletoes; Calder, D., Bernhardt, P., Eds.; Academic Press: London, UK, 1983. [Google Scholar]
- Bandaranayake, P.C.G.; Yoder, J.I. Haustorium Initiation and Early Development. In Parasitic Orobanchaceae: Parasitic Mechanisms and Control Strategies; Joel, D.M., Gressel, J., Musselman, L.J., Eds.; Springer: Berlin Heidelberg, Germany, 2013; pp. 61–74. [Google Scholar]
- Saucet, S.B.; Shirasu, K. Molecular Parasitic Plant-Host Interactions. PLoS Pathog. 2016, 12, e1005978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokla, A.; Melnyk, C.W. Developing a thief: Haustoria formation in parasitic plants. Dev. Biol. 2018, 442, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Keyes, W.J.; Palmer, A.G.; Erbil, W.K.; Taylor, J.V.; Apkarian, R.P.; Weeks, E.R.; Lynn, D.G. Semagenesis and the parasitic angiosperm Striga asiatica. Plant J. Cell Mol. Biol. 2007, 51, 707–716. [Google Scholar] [CrossRef]
- Smith, J.L.; De Moraes, C.M.; Mescher, M.C. Jasmonate- and salicylate-mediated plant defense responses to insect herbivores, pathogens and parasitic plants. Pest Manag. Sci. 2009, 65, 497–503. [Google Scholar] [CrossRef]
- Runyon, J.B.; Mescher, M.C.; De Moraes, C.M. Plant defenses against parasitic plants show similarities to those induced by herbivores and pathogens. Plant Signal. Behavior. 2010, 5, 929–931. [Google Scholar] [CrossRef] [Green Version]
- Cuevas-Reyes, P.; Pérez-López, G.; Maldonado-López, Y.; González-Rodríguez, A. Effects of herbivory and mistletoe infection by Psittacanthus calyculatus on nutritional quality and chemical defense of Quercus deserticola along Mexican forest fragments. Plant Ecol. 2017, 218, 687–697. [Google Scholar] [CrossRef]
- Hu, B.; Sakakibara, H.; Takebayashi, Y.; Peters, F.S.; Schumacher, J.; Eiblmeier, M.; Arab, L.; Kreuzwieser, J.; Polle, A.; Rennenberg, H. Mistletoe infestation mediates alteration of the phytohormone profile and anti-oxidative metabolism in bark and wood of its host Pinus sylvestris. Tree Physiol. 2017, 37, 676–691. [Google Scholar] [CrossRef] [Green Version]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallé, G.L.; Armillotta, A.; Frochot, H. Mechanisms of resistance of four cultivars of poplar against Viscum album L. In Proceedings of the 3rd International Symposium of Parasitic Weeds, Aleppo, Syria, 7–10 May 1984; Parker, C., Musselman, L.J., Polhill, R.M., Wilson, A.K., Eds.; [Google Scholar]
- Bhat, K.A.; Akhtar, S.; Dar, N.A.; Bhat, M.I.; Bhat, F.A.; Rizwan, R.; Horielov, O.; Krasylenko, Y. Mistletoe Eradicator—A Novel Tool for Simultaneous Mechanical and Chemical Control of Mistletoe. J. Vis. Exp. 2022, 181, e63455. [Google Scholar] [CrossRef]
- Schrader-Patton, C.; Grulke, N.; Bienz, C. Assessment of Ponderosa Pine Vigor Using Four-Band Aerial Imagery in South Central Oregon: Crown Objects to Landscapes. Forests 2021, 12, 612. [Google Scholar] [CrossRef]
- Barbedo, J. A Review on the Use of Unmanned Aerial Vehicles and Imaging Sensors for Monitoring and Assessing Plant Stresses. Drones 2019, 3, 40. [Google Scholar] [CrossRef] [Green Version]
- Duarte, A.; Borralho, N.; Cabral, P.; Caetano, M. Recent Advances in Forest Insect Pests and Diseases Monitoring Using UAV-Based Data: A Systematic Review. Forests 2022, 13, 911. [Google Scholar] [CrossRef]
- Thapa, S. Detection and Mapping of Incidence of Viscum album in Pinus sylvestris Forest of Southern French Alpe Using Satellite and Airborne Optical Imagery. Master’s Thesis, University of Twente, Enskode, The Netherlands, 2013. [Google Scholar]
- Barbosa, J.; Sebastián-González, E.; Asner, G.; Knapp, D.; Anderson, C.; Martin, R.; Dirzo, R. Hemiparasite-host plant interactions in a fragmented landscape assessed via imaging spectroscopy and LiDAR. Ecol. Appl. 2016, 26, 55–66. [Google Scholar] [CrossRef]
- Lawley, V.; Lewis, M.; Clarke, K.; Ostendorf, B. Site-based and remote sensing methods for monitoring indicators of vegetation condition: An Australian review. Ecol. Indic. 2016, 60, 1273–1283. [Google Scholar] [CrossRef] [Green Version]
- Windmuller-Campione, M.A.; Moser, R.L. Remote and Seasonal Field Detection of Eastern Spruce Dwarf Mistletoe in Northern Minnesota; Minnesota Forestry Research Notes; Department of Forest Resources, University of Minnesota: St. Paul, MN, USA, 2022; Volume 315. [Google Scholar]
- Mejia-Zuluaga, P.A.; Dozal, L.; Valdiviezo-N., J.C. Genetic Programming Approach for the Detection of Mistletoe Based on UAV Multispectral Imagery in the Conservation Area of Mexico City. Remote Sens. 2022, 14, 801. [Google Scholar] [CrossRef]
- Pernar, R.; Bajic, M.; Ancic, M.; Seletkovic, A.; Idzojtic, M. Detection of mistletoe in digital colour infrared images of infested fir trees. Period. Biol. 2007, 109, 67–75. [Google Scholar]
- Sabrina, F.; Sohail, S.; Thakur, S.; Azad, S.; Wasimi, S. Use of deep learning approach on UAV imagery to detect mistletoe infestation. In Proceedings of the 2020 IEEE Region 10 Symposium (TENSYMP), Dhaka, Bangladesh, 5–7 June 2020; pp. 556–559. [Google Scholar]
- Ančić, M.; Pernar, R.; Bajić, M.; Seletković, A.; Kolić, J. Detecting mistletoe infestation on Silver fir using hyperspectral images. Iforest-Biogeosci. For. 2014, 7, 85. [Google Scholar] [CrossRef] [Green Version]
- Maes, W.H.; Huete, A.R.; Avino, M.; Boer, M.M.; Dehaan, R.; Pendall, E.; Griebel, A.; Steppe, K. Can UAV-based infrared thermography be used to study plant-parasite interactions between mistletoe and eucalypt trees? Remote Sens. 2018, 10, 2062. [Google Scholar] [CrossRef] [Green Version]
- Xiong, D.; Huang, H.; Wang, Z.; Li, Z.; Tian, C. Assessment of dwarf mistletoe (Arceuthobium sichuanense) infection in spruce trees by using hyperspectral data. For. Pathol. 2021, 51, e12669. [Google Scholar] [CrossRef]
- Miraki, M.; Sohrabi, H.; Fatehi, P.; Kneubuehler, M. Detection of mistletoe infected trees using UAV high spatial resolution images. J. Plant Dis. Prot. 2021, 128, 1679–1689. [Google Scholar] [CrossRef]
- León-Bañuelos, L.A.; Endara-Agramont, A.R.; Gómez-Demetrio, W.; Martínez-García, C.G.; Gabino Nava-Bernal, E. Identification of Arceuthobium globosum using unmanned aerial vehicle images in a high mountain forest of central Mexico. J. For. Res. 2020, 31, 1759–1771. [Google Scholar] [CrossRef]
- Hamamouch, N.; Westwood, J.H.; Banner, I.; Cramer, C.L.; Gepstein, S.; Aly, R. A peptide from insects protects transgenic tobacco from a parasitic weed. Transgenic Res. 2005, 14, 227–236. [Google Scholar] [CrossRef]
- Aly, R.; Dina, P.; Guy, A. Expression of sarcotoxin IA gene via a root-specific tob promoter enhanced host resistance against parasitic weeds in tomato plants. Plant Cell Rep. 2006, 25, 297–303. [Google Scholar] [CrossRef]
- Aly, R.; Cholakh, H.; Joel, D.M.; Leibman, D.; Steinitz, B.; Zelcer, A.; Naglis, A.; Yarden, O.; Gal-On, A. Gene silencing of mannose 6-phosphate reductase in the parasitic weed Orobanche aegyptiaca through the production of homologous dsRNA sequences in the host plant. Plant Biotechnol. J. 2009, 7, 487–498. [Google Scholar] [CrossRef]
- Alakonya, A.; Kumar, R.; Koenig, D.; Kimura, S.; Townsley, B.; Runo, S.; Garces, H.M.; Kang, J.; Yanez, A.; David-Schwartz, R.; et al. Interspecific RNA interference of SHOOT MERISTEMLESS-like disrupts Cuscuta pentagona plant parasitism. Plant Cell 2012, 24, 3153–3166. [Google Scholar] [CrossRef] [Green Version]
- Stanivuković, Z.; Milenković, M.; Karadžić, D.; Mihajlović, L. The mistletoe (Viscum album L.)-a problem in fir (Abies alba Mill.) forests in Serbia and the Republic of Srpska [Bosnia and Herzegovina]. In Proceedings of the Serbian Forestry Congress, Belgrade, Serbia, 11–13 November 2010. [Google Scholar]
- Idžojtić, M.; Glavaš, M.; Zebec, M.; Pernar, R.; Kušan, Ž.; List, Đ.; Grahovac-Tremski, M. Intensity of Infection with Yellow Mistletoe and White-berried Mistletoe on the Area of the Forest Administrations Zagreb and Koprivnica. Šumarski List 2008, 132, 107–114. [Google Scholar]
- Struwe, I.; Gertsson, C.; Coulianos, C. Insects monophagous on mistletoe (Viscum album L.) newly discovered in Sweden: Cacopsylla visci (Curtis, 1835)(Hemiptera, Psyllidae) and Pinalitus viscicola (Puton, 1888) (Hemiptera, Miridae). Entomol. Tidskr. 2009, 130, 155–160. [Google Scholar]
- Parker, A. Growth of Wallrothiella arceuthobii on artificial media. Can. J. Bot. 1970, 48, 837–838. [Google Scholar] [CrossRef]
- Ramsfield, T.; Shamoun, S.; van der Kamp, B. Infection of Arceuthobium americanum by Colletotrichum gloeosporioides and its potential for inundative biological control. For. Pathol. 2005, 35, 332–338. [Google Scholar] [CrossRef]
- Askew, S.E.; Shamoun, S.F.; Van Der Kamp, B.J. An in vitro method for screening Colletotrichum gloeosporioides as a biological control agent for western hemlock dwarf mistletoe. For. Pathol. 2009, 39, 279–288. [Google Scholar] [CrossRef]
- Askew, S.E.; Shamoun, S.F.; van der Kamp, B.J. Assessment of Colletotrichum gloeosporioides as a biological control agent for management of hemlock dwarf mistletoe (Arceuthobium tsugense). For. Pathol. 2011, 41, 444–452. [Google Scholar] [CrossRef]
- Parmeter, J.; Hood, J.; Scharpf, R. Colletotrichum blight of dwarf mistletoe. Phytopathology 1959, 49, 812–815. [Google Scholar]
- Shamoun, S. Development of biological control strategy for management of dwarf mistletoes. In Proceedings of the 45th Western International Forest Disease Work Conference, Prince George, Victoria, BC, Canada, 15–19 September 1997; Sturrock, R., Ed.; Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 1997; pp. 36–42. [Google Scholar]
- Stojanović, S. The investigation of Sphaeropsis visci (Salm.) Sacc. and Colletotrichum gloeosporoides (Sacc.) Penz., parasite on European mistletoe (Viscum album ssp. typicum Beck). Zaštita Bilja 1989, 40, 493–503. [Google Scholar]
- Deeks, S.J.; Shamoun, S.F.; Punja, Z.K. Histopathology of callus and germinating seeds of Arceuthobium tsugense subsp. tsugense infected by Cylindrocarpon cylindroides and Colletotrichum gloeosporioides. Int. J. Plant Sci. 2002, 163, 765–773. [Google Scholar] [CrossRef]
- Hawksworth, F.G. Biological Control of the Mistletoes. In Proceedings of the Fifteenth Congress of the International Union of Forestry Research Organizations, Gainesville, FL, USA, 14–20 March 1971; Biological Control of Forest Diseases. Nordin, V.J., Ed.; Canadian Forestry Service: Ottawa, ON, Canada, 1972; pp. 83–92. [Google Scholar]
- Karadžić, D.; Lazarev, V.S.; Milenković, M. The most significant parasitic and saprophytic fungi on common mistletoe (Viscum album L) and their potential application in biocontrol. Glas. Šumarskog Fak. 2004, 89, 115–126. [Google Scholar] [CrossRef]
- Byler, J.; Cobb, F.W., Jr. The occurrence and pathogenicity of Nectria fuckeliana on dwarf mistletoe in California. Can. J. Bot. 1972, 50, 1162. [Google Scholar] [CrossRef]
- Varga, I.; Baltazár, T.; Pejchal, M. Optimisation of growing conditions of European mistletoe hyperparasitic fungus (phaeobotryosphaeria visci): Effect of different media and antibiotics/optimalizácia rastových podmienok Phaeobotryosphaeria visci, hyperparazitickej huby imela bieleho: Vplyv rôznych živných médií a antibiotík. Acta Hortic. Et Reg. 2013, 16, 44–47. [Google Scholar]
- Varga, I.; Baltazár, T.; Apró, M.; Poczai, P.; Hyvönen, J. Optimizing conditions for sporulation of European mistletoe hyperparasitic fungus (Phaeobotryosphaeria visci): Effect of light and different media. J. Agric. Sci. 2012, 50, 60–66. [Google Scholar]
- Brandenburger, W. Parasitische Pilze an Gefäßpflanzen in Europa; Gustav Fischer Verlag: Stuttgart, Germany, 1985. [Google Scholar]
- Chen, J.; Liu, X.; Jia, H.; Zhu, W. First report of leaf-spot disease caused by Sphaeropsis visci on Asian mistletoe [Viscum coloratum (Kom.) Nakai] in China. J. For. Res. 2018, 29, 1769–1774. [Google Scholar] [CrossRef]
- Haque, M.; Badshah, K. Loranthus parasitism-a challenge to the development of economic tree resource in the Rawalpindi East Region. Pak. J. For. 1984, 34, 101–109. [Google Scholar]
- Holecová, M.; Hollá, K.; Šebestová, M. Poznámky k rozšíreniu Ixapion Variegatum (Wencker, 1864) (Coleoptera, Curculionoidea, Apionidae) na území borskej nížiny (Jz Slovensko). Folia Faun. Slovaca 2016, 21, 9–12. [Google Scholar]
Figure 1.
Mistletoe parasitism and impacts on a tree host. Specific mistletoe may parasitize its host by exploiting its branch(es) for water and various other resources, thereby limiting its photosynthetic potential, optimal growth, response to various stresses, and fruit or timber production. Gradually, infective spread (by haustorial connections and endophytic expansion) and growing infection instances (ensured by mistletoe seed dispersal) may lead to early senescence of the tree(s).
Figure 1.
Mistletoe parasitism and impacts on a tree host. Specific mistletoe may parasitize its host by exploiting its branch(es) for water and various other resources, thereby limiting its photosynthetic potential, optimal growth, response to various stresses, and fruit or timber production. Gradually, infective spread (by haustorial connections and endophytic expansion) and growing infection instances (ensured by mistletoe seed dispersal) may lead to early senescence of the tree(s).

Figure 2.
A generalized life cycle of a mistletoe parasite on a tree host. Following seed dispersal, a sticky material, viscin, over the seed coat ensures firm attachment to the host branch (in 1) [87,88]. Hypocotyl(s) from the seed then emerges and grows towards the bark (in 2), which eventually, while attaching to the bark (in 3), penetrates through it (in 4) [89]. Once the hypocotyl connects to the host xylem (in 5), it exploits the host for water, minerals, carbohydrates, and secondary metabolites (in 6) [83]. The endophytic system may expand as sinkers (or epicortical roots) along the host cambium, from which sporadically new shoots may emerge (in 7) and facilitate the heightening of infectivity [65].
Figure 2.
A generalized life cycle of a mistletoe parasite on a tree host. Following seed dispersal, a sticky material, viscin, over the seed coat ensures firm attachment to the host branch (in 1) [87,88]. Hypocotyl(s) from the seed then emerges and grows towards the bark (in 2), which eventually, while attaching to the bark (in 3), penetrates through it (in 4) [89]. Once the hypocotyl connects to the host xylem (in 5), it exploits the host for water, minerals, carbohydrates, and secondary metabolites (in 6) [83]. The endophytic system may expand as sinkers (or epicortical roots) along the host cambium, from which sporadically new shoots may emerge (in 7) and facilitate the heightening of infectivity [65].

Figure 3.
Selective traits for testing and screening of potential MBCAs. The infographic depicts a generalized checklist of features desirable for testing and screening of putative MBCAs. A more detailed checklist should include features sought for MBCA formulations of specific organismic entities, viz., microbes, insects, etc. Some features in the above are adapted from known monographs [216,255].
Figure 3.
Selective traits for testing and screening of potential MBCAs. The infographic depicts a generalized checklist of features desirable for testing and screening of putative MBCAs. A more detailed checklist should include features sought for MBCA formulations of specific organismic entities, viz., microbes, insects, etc. Some features in the above are adapted from known monographs [216,255].

Figure 4.
Resource-tapping possibilities from genetic pools to strategize transgenic host resistance to mistletoes. The gap in functional genomics studies on mistletoe parasitism has majorly limited the exogenous induction and/or introduction of mistletoe resistance in hosts. This would require, in principle, the characterization of genetic targets in mistletoe and tree hosts that signal and favor easy entry to mistletoes (shown here) and then testing of the conventional gene manipulation and transgenics’ frameworks (see Figure 5). Even putative MBCAs can be screened for genetic elements encoding factors detrimental to mistletoe parasitism and/or its life cycle.
Figure 4.
Resource-tapping possibilities from genetic pools to strategize transgenic host resistance to mistletoes. The gap in functional genomics studies on mistletoe parasitism has majorly limited the exogenous induction and/or introduction of mistletoe resistance in hosts. This would require, in principle, the characterization of genetic targets in mistletoe and tree hosts that signal and favor easy entry to mistletoes (shown here) and then testing of the conventional gene manipulation and transgenics’ frameworks (see Figure 5). Even putative MBCAs can be screened for genetic elements encoding factors detrimental to mistletoe parasitism and/or its life cycle.

Figure 5.
Generalized transgenic and translational research pipeline for developing and disseminating stable mistletoe-resistant tree hosts of commercial value(s). Genetic elements (or their homologs) putatively conferring mistletoe resistance may be recovered from relevant repositories such as the NCBI’s GenBank and KEGG, and also may be screened using systems biology and bioinformatics tools, following genomic, transcriptomic, and metabolomic analyses. Some suggestions are depicted in Figure 4. Precisely, such meta-analyses can be performed on various pairs of genetic/expression pools. Once the genetic elements directly encoding the resistance trait or indirectly influencing the parasitism are stipulated, they may be tested on various grounds in small-scale assays and observations, in order to plan the appropriate design of the transgenic framework. In this way, cell and tissue culture models of parasitism involving trees and mistletoes may be indispensable and may generate proof(s) of concept(s). Then, a focused transgenic gene manipulation and characterization stage would involve lab-to-field-scale testing and validation of the successfully developed events. Translational activities would follow commercialization requisites on a case-by-case basis.
Figure 5.
Generalized transgenic and translational research pipeline for developing and disseminating stable mistletoe-resistant tree hosts of commercial value(s). Genetic elements (or their homologs) putatively conferring mistletoe resistance may be recovered from relevant repositories such as the NCBI’s GenBank and KEGG, and also may be screened using systems biology and bioinformatics tools, following genomic, transcriptomic, and metabolomic analyses. Some suggestions are depicted in Figure 4. Precisely, such meta-analyses can be performed on various pairs of genetic/expression pools. Once the genetic elements directly encoding the resistance trait or indirectly influencing the parasitism are stipulated, they may be tested on various grounds in small-scale assays and observations, in order to plan the appropriate design of the transgenic framework. In this way, cell and tissue culture models of parasitism involving trees and mistletoes may be indispensable and may generate proof(s) of concept(s). Then, a focused transgenic gene manipulation and characterization stage would involve lab-to-field-scale testing and validation of the successfully developed events. Translational activities would follow commercialization requisites on a case-by-case basis.

Figure 6.
Scheme of existing mistletoe management options and future possibilities with biotechnology and technologically smart approaches. Transgenic interventions and the use of technological advancements may solve the present problems in mistletoe management. An epigenetic remodeling strategy, however, would be a far-future possibility and might be realized following a full understanding of the basis of the multitrophic interactions prevailing in the vicinity or within mistletoe–tree host associations.
Figure 6.
Scheme of existing mistletoe management options and future possibilities with biotechnology and technologically smart approaches. Transgenic interventions and the use of technological advancements may solve the present problems in mistletoe management. An epigenetic remodeling strategy, however, would be a far-future possibility and might be realized following a full understanding of the basis of the multitrophic interactions prevailing in the vicinity or within mistletoe–tree host associations.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mudgal, G.; Kaur, J.; Chand, K.; Parashar, M.; Dhar, S.K.; Singh, G.B.; Gururani, M.A. Mitigating the Mistletoe Menace: Biotechnological and Smart Management Approaches. Biology 2022, 11, 1645. https://doi.org/10.3390/biology11111645
AMA Style
Mudgal G, Kaur J, Chand K, Parashar M, Dhar SK, Singh GB, Gururani MA. Mitigating the Mistletoe Menace: Biotechnological and Smart Management Approaches. Biology. 2022; 11(11):1645. https://doi.org/10.3390/biology11111645
Chicago/Turabian StyleMudgal, Gaurav, Jaspreet Kaur, Kartar Chand, Manisha Parashar, Sanjoy K. Dhar, Gajendra B. Singh, and Mayank A. Gururani. 2022. "Mitigating the Mistletoe Menace: Biotechnological and Smart Management Approaches" Biology 11, no. 11: 1645. https://doi.org/10.3390/biology11111645
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.