
Knowledge Gaps in Taxonomy, Ecology, Population Distribution Drivers and Genetic Diversity of African Sandalwood (Osyris lanceolata Hochst. & Steud.): A Scoping Review for Conservation

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Methodology
2.1. Definitions
2.2. Study Review Design
Search Process
3. Result of Search
3.1. Taxonomy of Osyris sp.
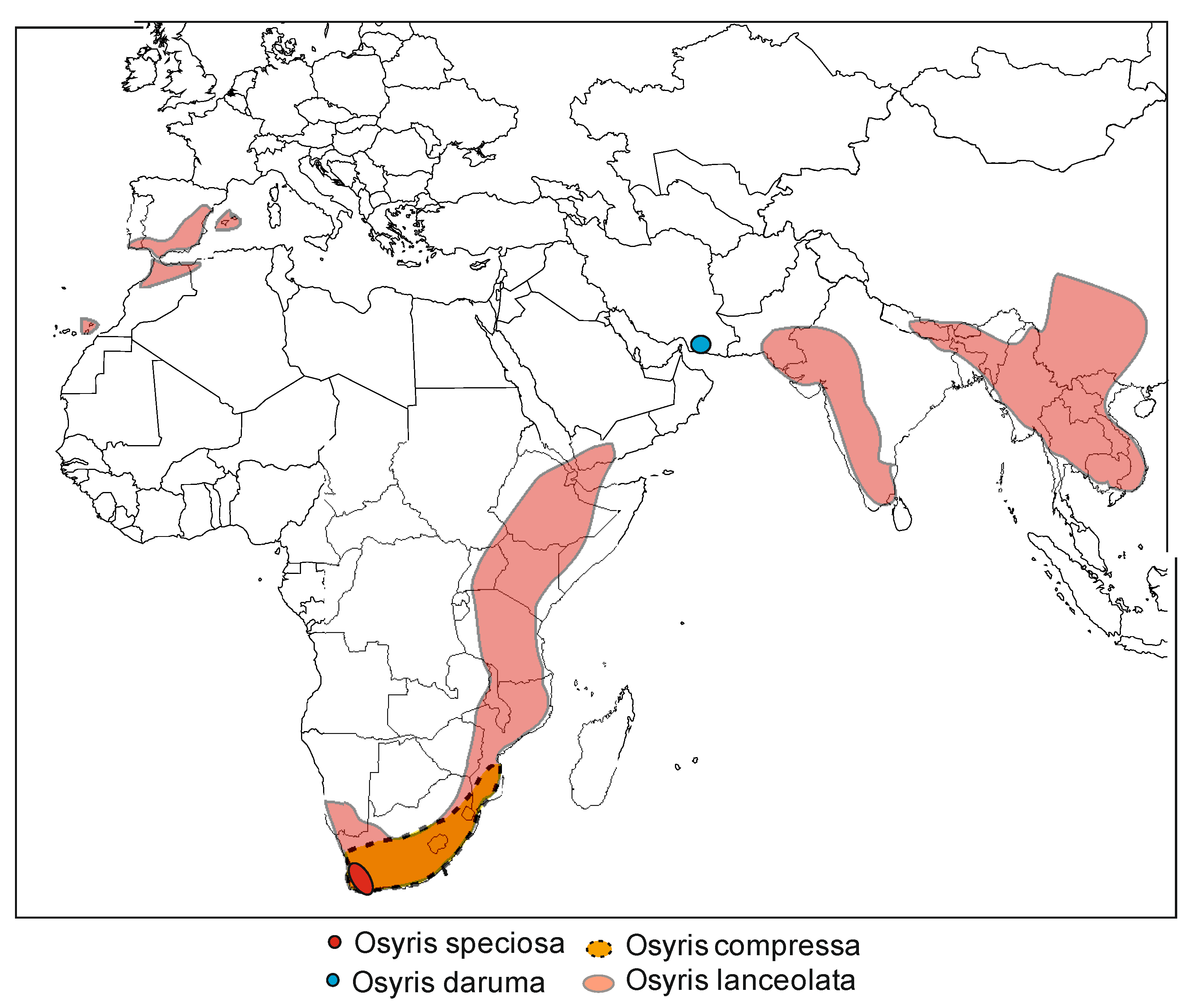
Global Distribution of Osyris lanceolata
3.2. Biology of African Sandalwood: Hemiparasitism and the Missing Links
Genetic Studies on African Sandalwood
3.3. Ecology: Habitats and Drivers for Distribution of African Sandalwood
3.3.1. Population Dynamics of Osyris lanceolata in Sub-Saharan Africa
3.3.2. Ethnobotany of African Sandalwood
3.4. Implications for Conservation of Osyris lanceolata in Sub-Saharan Africa
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Species | Synonym IPNI | Synonym COL | Synonym WFO | Synonym Tropicos |
|---|---|---|---|---|
| Osyris L. | ||||
| Osyris alba L. | ||||
| Osyris mediterranea Bubani | Osyris mediterranea Bubani | Osyris mediterranea Bubani | ||
| Osyris compressa (P.J. Bergius) A. DC. | ||||
| Osyris daruma Parsa | ||||
| Osyris speciosa (A.W. Hill) J.C. Manning & Goldblatt | ||||
| Osyris lanceolata Steud. & Hochst. ex A.DC. | ||||
| Osyris quadripartita Salzm. ex Decne. | Osyris quadripartita Salzm. ex Decne. | Osyris quadripartita Salzm. ex Decne. | ||
| Osyris abyssinica Hochst. ex A. Rich. | Osyris abyssinica Hochst. ex A. Rich. | Osyris abyssinica Hochst. ex A. Rich. | ||
| Osyris densifolia Peter, Fedde, Repert | Osyris densifolia Peter | Osyris densifolia Peter | ||
| Osyris laeta Peter, Fedde, Repert. | Osyris laeta Peter | Osyris laeta Peter | ||
| Osyris oblanceolate Peter | Osyris oblanceolate Peter | Osyris oblanceolate Peter | ||
| Osyris parvifolia Baker | Osyris parvifolia Baker | Osyris parvifolia Baker | ||
| Osyris pendula Balf.f | Osyris pendula Balf.f | |||
| Osyris quadrifida | ||||
| Osyris quadripartita var. | Osyris quadripartita var. puberula Kumari | |||
| Osyris rigidissima Engl. | Osyris rigidissima Engl. | Osyris rigidissima Engl. | ||
| Osyris tenuifolia Engl. | Osyris tenuifolia Engl. | Osyris tenuifolia Engl. | ||
| Osyris urundiensis De Wild | Osyris urundiensis De Wild. | Osyris urundiensis De Wild. | ||
| Osyris wightiana Wall. ex Wight | Osyris wightiana Wall. ex Wight | Osyris wightiana Wall. ex Wight | ||
| Osyris divaricata Pilg | ||||
| Osyris arborea Wall. | Osyris arborea wall. ex A. DC. | Osyris arborea wall. ex A. DC. | ||
| Osyris nepalensis Griff | Osyris nepalensis Griff | |||
| Osyris wightiana var. puberula (Hook.fil.) | ||||
| Osyris wightiana Wall. | Osyris wightiana var. rotundifolia (P.C. Tam) | Osyris wightiana var. rotundifolia (P.C. Tam) P.C. Tam | ||
| Osyris arborea var. rotundifolia P.C. Tam | Osyris arborea var. rotundifolia P.C. Tam | |||
| Osyris arborea var, spitata Lecomte | Osyris arborea var, spitata Lecomte | |||
| Osyris abyssinica var. speciosa A.W. Hill | Osyris abyssinica var. speciosa A.W. Hill | |||
| Osyris wightiana var. spitata (Lecomte) P.C. Tam | Osyris wightiana var wightiana | |||
| Osyris spinescens Mart. & Eichler | ||||
| Osyris peltate Roxb. | ||||
References
- da Silva, J.A.T.; Kher, M.M.; Soner, D.; Nataraj, M.; Dobránszki, J.; Millar, M.A. Santalum molecular biology: Molecular markers for genetic diversity, phylogenetics and taxonomy, and genetic transformation. Agrofor. Syst. 2018, 92, 1301–1315. [Google Scholar] [CrossRef]
- Ola, O.; Benjamin, E. Preserving Biodiversity and Ecosystem Services in West African Forest, Watersheds, and Wetlands: A Review of Incentives. Forests 2019, 10, 479. [Google Scholar] [CrossRef] [Green Version]
- Mbuya, L.P.; Msanga, H.P.; Ruffo, C.K.; Birnie, A.; Tengnas, B.O. Useful Trees and Shrubs for Tanzania; SIDA (Swedish International Development Authority): Nairobi, Kenya, 1994; 542p. [Google Scholar]
- Page, T.; Hannington, T.; Bunt, C.; Potrawiak, A.; Berry, A. Opportunities for the Smallholder Sandalwood Industry in Vanuatu; ACIAR Technical Reports No. 79; Australian Centre for International Agricultural Research: Canberra, Australia, 2012. [Google Scholar]
- Thomson, L.A.J. Looking ahead—Global sandalwood production and markets in 2040, and implications for Pacific Island producers. Aust. For. 2020, 83, 245–254. [Google Scholar] [CrossRef]
- Rao, M.N.; Ganeshaiah, K.N.; Shaanker, R.U. Assessing Threats and Mapping Sandal Resources to Identify Genetic ‘Hot-Spot’for In-Situ Conservation in Peninsular India. Conserv. Genet. 2007, 8, 925–935. [Google Scholar]
- Mwang’Ingo, P.L.; Teklehaimanot, Z.; Lulandala, L.L.; Mwihomeke, S.T. Host plants of Osyris lanceolata (African Sandalwood) and their influence on its early growth performance in Tanzania. S. Afr. For. J. 2005, 203, 55–65. [Google Scholar] [CrossRef]
- CITES. Consideration of Proposals for Amendment of Appendices I and II; CITES: Geneva, Switzerland, 2013; pp. 1–10. [Google Scholar]
- Muhoozi, J. Encroachers Target Kabaruka in Akagera. The East African. Available online: http://www.theeastafrican.co.ke/rwanda/News/Encroachers-target-Kabaruka-in-Akagera/1433218-2093116-sy2phc/index.html (accessed on 23 March 2018).
- Tajuba, P. How Oil Firm Is Burning Up Karamoja Valuable Trees. Available online: http://www.monitor.co.ug/artsculture/Reviews/oil-firm-burning-up-Karamoja-valuable-trees/691232-2996318-r5xq40/index.html (accessed on 6 August 2021).
- Bunei, E.K. The Hunt for the Precious Wood: Illegal Trade of Sandalwood as an International Criminal Enterprise in Kenya. Soc. Bus. Rev. 2017, 12, 63–76. [Google Scholar] [CrossRef]
- Wilson, B. Osyris lanceolata. The IUCN Red List of Threatened Species 2018: e.T200642A2675362. Available online: https://www.iucnredlist.org/species/200642/2675362 (accessed on 2 July 2021).
- USAID. Uganda Environmental Threats and Opportunities Assessment (Etoa) Final Etoa; USAID: Washington, DC, USA, 2015. [Google Scholar]
- Uganda Investment Authority (UIA). Karamoja Investment Profile; Uganda Investment Authority: Kampala, Uganda, 2016. [Google Scholar]
- Neuhauser, C.; Andow, D.A.; Heimpel, G.E.; May, G.; Shaw, R.G.; Wagenius, S. Community Genetics: Expanding the Synthesis of Ecology and Genetics. Ecology 2003, 84, 545–558. [Google Scholar] [CrossRef]
- Mwang’ingo, P.L.; Teklehaimanot, Z.; Hall, J.B.; Zilihona, J.E. Sex Distribution, Reproductive Biology and Regeneration in the Dioecious Species Osyris lanceolata (African Sandalwood) in Tanzania. Tanzan. J. For. Nat. Conserv. 2007, 76, 118–133. [Google Scholar]
- Kalisz, S.; Nason, J.D.; Hanzawa, F.M.; Tonsor, S.J. Spatial Population Genetic Structure in Trillium Grandiflorum: The Roles of Dispersal, Mating, History, and Selection. Evolution 2001, 55, 1560–1568. [Google Scholar] [CrossRef]
- Curto, M.; Nogueira, M.; Beja, P.; Amorim, F.; Schümann, M.; Meimberg, H. Influence of past agricultural fragmentation to the genetic structure of Juniperus oxycedrus in a Mediterranean landscape. Tree Genet. Genomes 2015, 11, 11. [Google Scholar] [CrossRef]
- Tabuti, J.R.S.; Mugula, B.B. The ethnobotany and ecological status of Albizia coriaria Welw. Ex Oliv. In Budondo Sub-county, eastern Uganda. Afr. J. Ecol. 2007, 45, 126–129. [Google Scholar] [CrossRef]
- Virillo, C.B.; Martins, F.R.; Tamashiro, J.Y.; Dos Santos, F.A.M. Is size structure a good measure of future trends of plant populations? An empirical approach using five woody species from the Cerrado (Brazilian savanna). Acta Bot. Bras. 2011, 25, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Muchugi, A.; Lengkeek, A.G.; Kadu, C.A.C.; Muluvi, G.M.; Njagi, E.N.M.; Dawson, I.K. Genetic variation in the threatened medicinal tree Prunus africana in Cameroon and Kenya: Implications for current management and evolutionary history. S. Afr. J. Bot. 2006, 72, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Kadu, C.A.C.; Parich, A.; Schueler, S.; Konrad, H.; Muluvi, G.M.; Eyog-Matig, O.; Muchugi, A.; Williams, V.; Ramamonjisoa, L.; Kapinga, C.; et al. Bioactive constituents in Prunus africana: Geographical variation throughout Africa and associations with environmental and genetic parameters. Phytochemistry 2012, 83, 70–78. [Google Scholar] [CrossRef]
- Qasem, J.R. Host range of the parasitic weed Osyris alba L. in Jordan. Weed Biol. Manag. 2006, 6, 74–78. [Google Scholar] [CrossRef]
- Nurochman, D.; Matangaran, J.R.; Santosa, G.; Suharjito, D.; Sari, R.K. Autecology and morphological properties of sandalwood (Santalum album) in Pidie District, Aceh, Indonesia. Biodivers. J. Biol. Divers. 2018, 19, 406–412. [Google Scholar] [CrossRef]
- Gathara, M.; Makenzi, P.; Kimondo, J.; Muturi, G. Prediction of Osyris lanceolata (Hochst. Steud.) Site Suitability Using Indicator Plant Species and Edaphic Factors in Humid Highland and Dry Lowland Forests in Kenya. J. Hortic. For. 2014, 6, 99–106. [Google Scholar]
- Munn, Z.; Peters, M.D.J.; Stern, C.; Tufanaru, C.; McArthur, A.; Aromataris, E. Systematic review or scoping review? Guidance for authors when choosing between a systematic or scoping review approach. BMC Med. Res. Methodol. 2018, 18, 143. [Google Scholar] [CrossRef]
- Arksey, H.; O’Malley, L. Scoping studies: Towards a methodological framework. Int. J. Soc. Res. Methodol. 2005, 8, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Lauwers, L.; Bastiaens, H.; Remmen, R.; Keune, H. Nature’s Contributions to Human Health: A Missing Link to Primary Health Care? A Scoping Review of International Overview Reports and Scientific Evidence. Front. Public Health 2020, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Osyris lanceolata Hochst. & Steud. In GBIF Secretariat. 2021. Available online: https://doi.org/10.15468/39omei (accessed on 25 June 2021).
- The International Plant Names Index Collaborators. International Plant Names Index. 2019. Available online: https://doi.org/10.15468/uhllmw (accessed on 25 June 2021).
- Mwang’ingo, P.L.; Teklehaimanot, Z.; Hall, J.B.; Lulandala, L.L.L. African Sandalwood (Osyris lanceolata): Resource Assessment and Quality Variation among Populations in Tanzania: Research Note. S. Afr. For. J. 2003, 199, 77–88. [Google Scholar] [CrossRef]
- Masevhe, N.A.; McGaw, L.J.; Eloff, J.N. The traditional use of plants to manage candidiasis and related infections in Venda, South Africa. J. Ethnopharmacol. 2015, 168, 364–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, K.V.; Balasundaran, M.; Balagopalan, M. Identification of Santalum Album and Osyris Lanceolata through Morphological and Biochemical Characteristics and Molecular Markers to Check Adulteration; Kerala Forest Research Institute: Thrissur, India, 2006. [Google Scholar]
- Andiego, K.P.; Dangasuk, O.G.; Odee, D.W.; Omondi, F.S.; Otieno, D.F.; Balozi, B.K. Genetic diversity of endangered sandalwood (Osyris lanceolata) populations in Kenya using ISSR molecular markers. East Afr. Agric. For. J. 2019, 83, 80–93. [Google Scholar] [CrossRef]
- Otieno, J.O.; Omondi, S.F.; Perry, A.; Odee, D.W.; Makatiani, E.T.; Kiplagat, O.; Cavers, S. Development and characterization of microsatellite markers for Osyris lanceolata Hochst. & Steud., an endangered African sandalwood tree species. Trop. Plant Res. 2016, 3, 701–703. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.; Borger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwang’Ingo, P.L.; Teklehaimanot, Z.; Maliondo, S.M.; Msanga, H.P. Storage and pre-sowing treatment of recalcitrant seeds of Africa sandalwood (Osyris lanceolata). Seed Sci. Technol. 2004, 32, 547–560. [Google Scholar] [CrossRef]
- Wambua, J.K. The Distribution, Abundance and Ecological Impacts of Invasive Plant Species at Ol-Donyo Sabuk National Park, Kenya. Ph.D. Thesis, University of Nairobi, Nairobi, Kenya, 2010. [Google Scholar]
- Erbo, K.; Tolera, M.; Awas, T. Distribution, Association and Population Structure of Osyris Quadripartita (African Sandalwood) in a Dry Woodland Forest, Southern Ethiopia. Glob. J. Biol. Agric. Health Sci. 2020, 9, 101. [Google Scholar] [CrossRef]
- Abascal, F.; Corvelo, A.; Cruz, F.; Villanueva-Cañas, J.L.; Vlasova, A.; Marcet-Houben, M.; Martínez-Cruz, B.; Cheng, J.; Prieto, P.; Quesada, V.; et al. Extreme genomic erosion after recurrent demographic bottlenecks in the highly endangered Iberian lynx. Genome Biol. 2016, 17, 251. [Google Scholar] [CrossRef] [Green Version]
- Teklehaimanot, Z.; Mwang’Ingo, P.L.; Mugasha, A.G.; Ruffo, C.K. Influence of the origin of stem cutting, season of collection and auxin application on the vegetative propagation af African Sandalwood (Osyris lanceolata) in Tanzania. S. Afr. For. J. 2004, 201, 13–24. [Google Scholar] [CrossRef]
- Kamondo, B.; Juma, B.; Mwangi, L.; Meroka, D. Domestication of Osyris lanceolata in Kenya: Propagation, Conservation, Management and Commercialisation; Annual Report; Muguga Regional Research Centre: Nairobi, Kenya, 2007. [Google Scholar]
- Osyris lanceolata Hochst. Osyris lanceolata Hochst. & Steud. In The Plant List 2016. Available online: http://www.theplantlist.org/tpl1.1/record/kew-2396402 (accessed on 6 August 2021).
- Polhill, P.M. Flora of Tropical East Africa: Santalaceae; Royal Botanic Gardens Kew: Richmond, UK, 2005. [Google Scholar]
- Nickrent, D. Status of the Genera Colpoon, Osyris and Rhoiacarpos in South Africa Molecular Phylogenetic Data. No. 1961. Bothalia Afr. Biodivers. Conserv. 2017, 47, 1–7. [Google Scholar]
- Fay, M.F.; Bennett, J.R.; Dixon, K.W.; Christenhusz, M.J.M. Parasites, their relationships and the disintegration of Scrophulariaceae sensu lato. Curtis’s Bot. Mag. 2010, 26, 286–313. [Google Scholar] [CrossRef]
- Mugula, B.B. Field Surveys for Mapping Osyris Lanceolata in Karamoja, Sub-Region, Uganda. Field Work; 2020; (Unpublished). [Google Scholar]
- Breitenbach, F. The Indigenous Trees of Ethiopia, 2nd ed.; Ethiopian Forestry Association: Addis Ababa, Ethiopian, 1963. [Google Scholar]
- Bahadur, K.K. Current status, distribution pattern and associations of Sandalwood (Santalum album) in Pyuthan District of Nepal. Clar. Int. Multidiscip. J. 2018, 7, 1–11. [Google Scholar] [CrossRef]
- Harbaugh, D.T.; Baldwin, B.G. Phylogeny and biogeography of the sandalwoods (Santalum, Santalaceae): Repeated dispersals throughout the Pacific. Am. J. Bot. 2007, 94, 1028–1040. [Google Scholar] [CrossRef] [PubMed]
- Ndangalasi, H.J.; Mligo, C.; Mvungi, E.F. Composition and Size Class Structure of Tree Species in Ihang’ana Forest Reserve, Mufindi District, Tanzania. Tanzan. J. Sci. 2014, 40, 1–12. [Google Scholar]
- Schueler, L.; Hemp, A. Atlas of Pollen and Spores and Their Parent Taxa of Mt Kilimanjaro and Tropical East Africa. Quat. Int. 2016, 425, 301–386. [Google Scholar] [CrossRef]
- Report on Africa Regional Meeting, CITES Tree Species Programme (CTSP) 10–15 March 2019, Dar Es Salaam, Tanzania. 2019, Volume 62, pp. 14–16. Available online: https://cites-tsp.org/wp-content/uploads/2020/02/CITES-Tree-Species-Programme-Regional-Meeting-for-Africa-11-to-15-March-2019-Dar-es-Salaam-Tanzania-%E2%80%93-Minutes..pdf (accessed on 20 August 2021).
- Ochanda, K.V. Conservation and Management of Sandalwood Trees: (Osyris lanceolata Hochst & Steudel,) in Chyullu Hills Kibwezi District, Kenya. MSc. Thesis, Kenyatta University, Nairobi, Kenya, 2009. [Google Scholar]
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Simons, A. Agroforestree Database: A Tree Reference and Selection Guide. Version 4; World Agroforestry Centre Nairobi Kenya: Nairobi, Kenya, 2009. [Google Scholar]
- Turini, F.G.; Steinert, C.; Heubl, G.; Bringmann, G.; Lombe, B.K.; Mudogo, V.; Meimberg, H. Microsatellites facilitate species delimitation in Congolese Ancistrocladus (Ancistrocladaceae), a genus with pharmacologically potent naphthylisoquinoline alkaloids. Taxon 2014, 63, 329–341. [Google Scholar] [CrossRef]
- Meimberg, H.; Rischer, H.; Turini, F.G.; Chamchumroon, V.; Dreyer, M.; Sommaro, M.; Bringmann, G.; Heubl, G. Evidence for species differentiation within the Ancistrocladus tectorius complex (Ancistrocladaceae) in Southeast Asia: A molecular approach. Plant Syst. Evol. 2010, 284, 77–98. [Google Scholar] [CrossRef]
- Perrino, E.V.; Silletti, G.N.; Erben, M.; Wagensommer, R.P. Viola Cassinensis Subsp. Lucana (Violaceae), a New Subspecies from the Lucanian Apennine, Southern Italy. Phyton Ann. Rei Bot. 2018, 58, 109–115. [Google Scholar]
- Kokwaro, J.O. Osyris lanceolata. In Medicinal Plants of East Africa, 3rd ed.; University of Nairobi Press: Nairobi, Kenya, 2009. [Google Scholar]
- Tennakoon, K.U.; Cameron, D.D. The anatomy of Santalum album (Sandalwood) haustoria. Can. J. Bot. 2006, 84, 1608–1616. [Google Scholar] [CrossRef]
- Okubamichael, D.Y.; Griffiths, M.E.; Ward, D. Host specificity in parasitic plants—Perspectives from mistletoes. AoB Plants 2016, 8, 8. [Google Scholar] [CrossRef]
- Bell, T.L.; Adams, M. Attack on all fronts: Functional relationships between aerial and root parasitic plants and their woody hosts and consequences for ecosystems. Tree Physiol. 2011, 31, 3–15. [Google Scholar] [CrossRef]
- Marvier, M.A. Parasitic Plant-Host Interactions: Plant Performance and Indirect Effects on Parasite-Feeding Herbivores. Ecology 1996, 77, 1398–1409. [Google Scholar] [CrossRef]
- Nge, F.J.; Ranathunge, K.; Kotula, L.; Cawthray, G.; Lambers, H. Strong host specificity of a root hemi-parasite (Santalum acuminatum) limits its local distribution: Beggars can be choosers. Plant Soil 2019, 437, 159–177. [Google Scholar] [CrossRef]
- Furuhashi, T.; Furuhashi, K.; Weckwerth, W. The parasitic mechanism of the holostemparasitic plant Cuscuta. J. Plant Interact. 2011, 6, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Kuijt, J. The Biology of Parasitic Flowering Plants; University of California Press: Berkeley, CA, USA, 1969. [Google Scholar]
- Watson, D.M. Determinants of parasitic plant distribution: The role of host quality. Botany 2009, 87, 16–21. [Google Scholar] [CrossRef]
- Irving, L.J.; Cameron, D.D. You are What You Eat: Interactions between Root Parasitic Plants and Their Hosts. Adv. Bot. Res. 2009, 50, 87–138. [Google Scholar]
- Yoder, J.I. Parasitic plant responses to host plant signals: A model for subterranean plant–plant interactions. Curr. Opin. Plant Biol. 1999, 2, 65–70. [Google Scholar] [CrossRef]
- Westwood, J.H.; Yoder, J.I.; Timko, M.P.; Depamphilis, C.W. The evolution of parasitism in plants. Trends Plant Sci. 2010, 15, 227–235. [Google Scholar] [CrossRef]
- Tomilov, A.; Tomilova, N.; Shin, D.H.; Jamison, D.; Torres, M.; Reagan, R.; Mcgray, H.; Horning, T.; Truong, R.; Nava, A.J.; et al. Chemical Signalling between Plants. In Chemical Ecology: From Gene to Ecosystem; Springer: Dordrecht, The Netherlands, 2006; pp. 55–69. [Google Scholar]
- Nilsson, C.H.; Svensson, B.M. Host affiliation in two subarctic hemiparasitic plants: Bartsia alpina and Pedicularis lapponica. Écoscience 1997, 4, 80–85. [Google Scholar] [CrossRef]
- Matthies, D.; Egli, P. Response of a root hemiparasite to elevated CO2 depends on host type and soil nutrients. Oecologia 1999, 120, 156–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tennakoon, K.U.; Pate, J.S. Heterotrophic gain of carbon from hosts by the xylem-tapping root hemiparasite Olax phyllanthi (Olacaceae). Oecologia 1996, 105, 369–376. [Google Scholar] [CrossRef]
- Vellend, M. Species Diversity and Genetic Diversity: Parallel Processes and Correlated Patterns. Am. Nat. 2005, 166, 199–215. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.L.; Burt, M.L.; Rexstad, E.; Thomas, L. Spatial models for distance sampling data: Recent developments and future directions. Methods Ecol. Evol. 2013, 4, 1001–1010. [Google Scholar] [CrossRef] [Green Version]
- Gacheri, N.; Wanjala, B.W.; Jamnadass, R.; Muchugi, A. Analysis of the Impact of Domestication of Warburgia ugandensis (Sprague) on Its Genetic Diversity Based on Amplified Fragment Length Polymorphism. Afr. J. Biotechnol. 2016, 15, 1673–1680. [Google Scholar]
- Ellegren, H.; Galtier, N. Determinants of genetic diversity. Nat. Rev. Genet. 2016, 17, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Indrioko, S.; Ratnaningrum, Y.W. Habitat Loss Caused Clonality, Genetic Diversity Reduction and Reproductive Failure in Santalum album (Santalaceae), an Endangered Endemic Species of Indonesia. Procedia Environ. Sci. 2015, 28, 657–664. [Google Scholar] [CrossRef] [Green Version]
- Farwig, N.; Braun, C.; Böhning-Gaese, K. Human disturbance reduces genetic diversity of an endangered tropical tree, Prunus africana (Rosaceae). Conserv. Genet. 2007, 9, 317–326. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Yao, J.; Li, Q.; Liu, F.; Yotsukura, N.; Krupnova, T.N.; Duan, D. Effect of domestication on the genetic diversity and structure of Saccharina japonica populations in China. Sci. Rep. 2017, 7, 42158. [Google Scholar] [CrossRef] [Green Version]
- Graudal, L.; Aravanopoulos, F.; Bennadji, Z.; Changtragoon, S.; Fady, B.; Kjaer, E.; Loo, J.; Ramamonjisoa, L.; Vendramin, G.G. Global to local genetic diversity indicators of evolutionary potential in tree species within and outside forests. For. Ecol. Manag. 2014, 333, 35–51. [Google Scholar] [CrossRef]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Yang, H.; Zhang, R.; Jin, G.; Feng, Z.; Zhou, Z. Assessing the Genetic Diversity and Genealogical Reconstruction of Cypress (Cupressus funebris Endl.) Breeding Parents Using SSR Markers. Forests 2016, 7, 160. [Google Scholar] [CrossRef] [Green Version]
- Vekemans, X.; Hardy, O. New insights from fine-scale spatial genetic structure analyses in plant populations. Mol. Ecol. 2004, 13, 921–935. [Google Scholar] [CrossRef]
- Petkova, D.; Novembre, J.; Stephens, M. Visualizing spatial population structure with estimated effective migration surfaces. Nat. Genet. 2016, 48, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Volis, S.; Ormanbekova, D.; Shulgina, I. Fine-scale spatial genetic structure in predominantly selfing plants with limited seed dispersal: A rule or exception? Plant Divers. 2016, 38, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQuillan, R.; Leutenegger, A.-L.; Abdel-Rahman, R.; Franklin, C.S.; Pericic, M.; Barac-Lauc, L.; Smolej-Narancic, N.; Janicijevic, B.; Polasek, O.; Tenesa, A.; et al. Runs of Homozygosity in European Populations. Am. J. Hum. Genet. 2008, 83, 359–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczecińska, M.; Sramko, G.; Wołosz, K.; Sawicki, J. Genetic Diversity and Population Structure of the Rare and Endangered Plant Species Pulsatilla patens (L.) Mill in East Central Europe. PLoS ONE 2016, 11, e0151730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porth, I.; El-Kassaby, Y.A. Assessment of the Genetic Diversity in Forest Tree Populations Using Molecular Markers. Diversity 2014, 6, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Holliday, J.A.; Aitken, S.N.; Cooke, J.E.K.; Fady, B.; Gonzalez-Martinez, S.C.; Heuertz, M.; Jaramillo-Correa, J.-P.; Lexer, C.; Staton, M.; Whetten, R.W.; et al. Advances in ecological genomics in forest trees and applications to genetic resources conservation and breeding. Mol. Ecol. 2016, 26, 706–717. [Google Scholar] [CrossRef]
- Castoe, T.A.; Poole, A.W.; de Koning, A.J.; Jones, K.L.; Tomback, D.F.; Oyler-McCance, S.; Fike, J.A.; Lance, S.; Streicher, J.; Smith, E.N.; et al. Rapid Microsatellite Identification from Illumina Paired-End Genomic Sequencing in Two Birds and a Snake. PLoS ONE 2012, 7, e30953. [Google Scholar] [CrossRef] [Green Version]
- De Barba, M.; Miquel, C.; Lobréaux, S.; Quenette, P.Y.; Swenson, J.E.; Taberlet, P. High-throughput microsatellite genotyping in ecology: Improved accuracy, efficiency, standardization and success with low-quantity and degraded DNA. Mol. Ecol. Resour. 2017, 17, 492–507. [Google Scholar] [CrossRef]
- Neophytou, C.; Torutaeva, E.; Winter, S.; Meimberg, H.; Hasenauer, H.; Curto, M. Analysis of microsatellite loci in tree of heaven (Ailanthus altissima (Mill.) Swingle) using SSR-GBS. Tree Genet. Genomes 2018, 14, 82. [Google Scholar] [CrossRef] [Green Version]
- Tibihika, P.D.; Curto, M.; Dornstauder-Schrammel, E.; Winter, S.; Alemayehu, E.; Waidbacher, H.; Meimberg, H. Application of microsatellite genotyping by sequencing (SSR-GBS) to measure genetic diversity of the East African Oreochromis niloticus. Conserv. Genet. 2019, 20, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Vartia, S.; Villanueva-Cañas, J.L.; Finarelli, J.; Farrell, E.D.; Collins, P.C.; Hughes, G.; Carlsson, J.E.L.; Gauthier, D.T.; McGinnity, P.; Cross, T.F.; et al. A novel method of microsatellite genotyping-by-sequencing using individual combinatorial barcoding. R. Soc. Open Sci. 2016, 3, 150565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šarhanová, P.; Pfanzelt, S.; Brandt, R.; Himmelbach, A.; Blattner, F.R. SSR-seq: Genotyping of microsatellites using next-generation sequencing reveals higher level of polymorphism as compared to traditional fragment size scoring. Ecol. Evol. 2018, 8, 10817–10833. [Google Scholar] [CrossRef] [PubMed]
- Parchman, T.L.; Jahner, J.P.; Uckele, K.A.; Galland, L.M.; Eckert, A.J. RADseq approaches and applications for forest tree genetics. Tree Genet. Genomes 2018, 14, 39. [Google Scholar] [CrossRef]
- Campbell, N.R.; Harmon, S.A.; Narum, S.R. Genotyping-in-Thousands by sequencing (GT-seq): A cost effective SNP genotyping method based on custom amplicon sequencing. Mol. Ecol. Resour. 2015, 15, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Dennenmoser, S.; Vamosi, S.M.; Nolte, A.W.; Rogers, S.M. Adaptive genomic divergence under high gene flow between freshwater and brackish-water ecotypes of prickly sculpin (Cottus asper) revealed by Pool-Seq. Mol. Ecol. 2017, 26, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.E.D. Why is Santalum Spicatum Common Near Granite Rocks? J. R. Soc. West. Aust. 1997, 80, 209. [Google Scholar]
- Pfenninger, M.; Salinger, M.; Haun, T.; Feldmeyer, B. Factors and processes shaping the population structure and distribution of genetic variation across the species range of the freshwater snail radix balthica (Pulmonata, Basommatophora). BMC Evol. Biol. 2011, 11, 135. [Google Scholar] [CrossRef] [Green Version]
- Amundrud, S.L. Abiotic and Biotic Processes Shape Species Distributions and Ecological Communities across Spatial Scales. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2020. [Google Scholar]
- Kamondo, B.; Giathi, G.; Osore, C.; Machua, J.; Kagunyu, L.; Wafula, A.; Bala, P.; Njuguna, J.; Wakori, S.; Maingi, F.; et al. Growing of East African Sandalwood: Guidelines for Tree Growers; Kenya Forestry Research Institute (KEFRI): Nairobi, Kenya, 2014. [Google Scholar]
- Mortelliti, A.; Amori, G.; Boitani, L. The role of habitat quality in fragmented landscapes: A conceptual overview and prospectus for future research. Oecologia 2010, 163, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F. Habitat destruction: Death by a thousand cuts. Conserv. Biol. All 2010, 1, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Chase, J.M.; Blowes, S.A.; Knight, T.M.; Gerstner, K.; May, F. Ecosystem decay exacerbates biodiversity loss with habitat loss. Nature 2020, 584, 238–243. [Google Scholar] [CrossRef]
- McKinnell, F.H.; Levinson, J. WA Sandalwood Industry Development Plan 2008–2020; Forest Products Commission: Kensington, Australia, 2008. [Google Scholar]
- Okiror, P.; Chono, J.; Nyamukuru, A.; Lwanga, J.S.; Sasira, P.; Diogo, P. Variation in Woody Species Abundance and Distribution in and around Kibale National Park, Uganda. Int. Sch. Res. Not. For. 2012, 2012, 490461. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, K.; Jeltsch, F.; Ward, D. Analysis of the population dynamics of Acacia trees in the Negev desert, Israel with a spatially-explicit computer simulation model. Ecol. Model. 1999, 117, 203–224. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Laurance, W.F. The ecology, distribution, conservation and management of large old trees. Biol. Rev. 2016, 92, 1434–1458. [Google Scholar] [CrossRef] [PubMed]
- Shyaula, S. A review on genus Osyris: Phytochemical constituents and traditional uses. J. Nat. Pharm. 2012, 3, 61. [Google Scholar] [CrossRef]
- Njoroge, G.N.; Bussmann, R.W. Diversity and utilization of antimalarial ethnophytotherapeutic remedies among the Kikuyus (Central Kenya). J. Ethnobiol. Ethnomed. 2006, 2, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moy, R.L.; Levenson, C. Sandalwood Album Oil as a Botanical Therapeutic in Dermatology. J. Clin. Aesthet. Dermatol. 2017, 10, 34–39. [Google Scholar]
- Dwivedi, C.; Abu-Ghazaleh, A. Chemopreventive effects of sandalwood oil on skin papillomas in mice. Eur. J. Cancer Prev. 1997, 6, 399–401. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, Y.; Khan, S.; Duan, G.; Chen, A.; Ling, L.; Zhao, L.; Liu, Z.; Wu, X. Accumulation of Pb, Cu, and Zn in native plants growing on contaminated sites and their potential accumulation capacity in Heqing, Yunnan. J. Environ. Sci. 2008, 20, 1469–1474. [Google Scholar] [CrossRef]
- Jain, R.; Nair, S. Sandalwood Oil for the Chemoprevention of Skin Cancer: Mechanistic Insights, Anti-inflammatory, and In Vivo Anticancer Potential. Curr. Pharmacol. Rep. 2019, 5, 345–358. [Google Scholar] [CrossRef]
- Coppen, J. Flavours and Fragrances of Plant Origin; Food and Agriculture Organization of the United Nations: Rome, Italy, 1995. [Google Scholar]
- Hemp, A.; Hemp, C.; Winter, J.C. Environment and Worldview: The Chagga Homegardens; Archaeopress: Oxford, UK, 2009. [Google Scholar]
- Bhowmik, D.; Biswas, D.; Kumar, K.P.S. Recent Aspect of Ethnobotanical Application and Medicinal Properties of Traditional Indian Herbs Santalum album. Int. J. Chem. Res. 2011, 1, 21–27. [Google Scholar]
- Akbar, S. Santalum album L. (Santalaceae). In Handbook of 200 Medicinal Plants; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1601–1608. [Google Scholar]
- Wang, C.; Kim, S.-W. Shaking up ancient scents: Insights into santalol synthesis in engineered Escherichia coli. Process. Biochem. 2015, 50, 1177–1183. [Google Scholar] [CrossRef]
- Cogoni, D.; Fenu, G.; Dessì, C.; Deidda, A.; Giotta, C.; Piccitto, M.; Bacchetta, G. Importance of Plants with Extremely Small Populations (PSESPs) in Endemic-Rich Areas, Elements Often Forgotten in Conservation Strategies. Plants 2021, 10, 1504. [Google Scholar] [CrossRef]
- Sulis, E.; Bacchetta, G.; Cogoni, D.; Fenu, G. From Global to Local Scale: Where is the Best for Conservation Purpose? Biodivers. Conserv. 2021, 30, 183–200. [Google Scholar] [CrossRef]
- Fenu, G.; Bacchetta, G.; Christodoulou, C.S.; Cogoni, D.; Fournaraki, C.; Gian Pietro, G.D.G.; Gotsiou, P.; Kyratzis, A.; Piazza, C.; Vicens, M.; et al. A Common Approach to the Conservation of Threatened Island Vascular Plants: First Results in the Mediterranean Basin. Diversity 2020, 12, 157. [Google Scholar] [CrossRef] [Green Version]
- WFO. Osyris lanceolata Hochst. & Steud. 2021. Available online: http://www.worldfloraonline.org/taxon/wfo-0000388245 (accessed on 25 July 2021).
- Missouri Botanical Garden. Available online: https://tropicos.org (accessed on 26 July 2021).


| Search Topic | Google Scholar | Web of Science | Scopus | Screened Papers | Author(s) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Santalum sp. | Osyris sp. | Santalum sp. | Osyris sp. | Santalum sp. | Osyris sp. | General Papers | Santalum sp. | Osyris sp. | Osyris sp. | |
| Population dynamics | 4760 | 833 | 2250 | 11 | 347 | 50 | 17 | 07 | 06 | [7,8,25,29,30,31] |
| Ethnobotanical uses | - | 06 | 01 | 71 | 18 | 16 | 11 | 05 | [3,29,30,31,32] | |
| Genetic diversity | 304.8 | 538 | 70 | 02 | 101 | 15 | 43 | 07 | 03 | [33,34,35] |
| Hemiparasitism | - | - | 01 | 00 | 22 | 04 | 03 | [24,36,37] | ||
| Distribution drivers | - | - | 00 | 01 | - | - | 09 | - | 03 | [38,39,40] |
| Propagation methods | - | - | 44 | 11 | - | - | 02 | [41,42] | ||
| Reproductive biology | 4030 | 01 | 01 | 393 | - | - | 01 | [16] | ||
| Species taxonomy | - | - | 12 | 01 | 125 | 32 | - | 05 | [29,30,43,44,45] | |
| African Region | Country Ranges | Local Populations (Reported) |
|---|---|---|
| East Africa | Uganda, Kenya, Tanzania, Rwanda, Burundi, South Sudan | Uganda: Karamoja subregion, Mbale (Kaburorun), West Nile, Bukwo, Kween and Kapchorwa [8,44,47] Kenya: Turkana, Baringo, Bogoria, Narok, Amboseli, Pokot, Samburu, Laikipia, Kajiado, Kitui, Taita hills, Chyulu hills, Gwasi hills, Marsabit, Makueni, Kikuyu escarpment forest, Mbeere, Narok, Ol-donyo Sabul, Oloitokitok and Mt. Kulal [8,38,44,52,53,54,55] Tanzania: Ufipa, Mbulu district, Mbisi, Lake Manyara, Songea, Ihang’ana forest, Kilimanjaro region, Masai Boma, Oldoinyo Sambu [7,30,41,44,52] Rwanda: Akagera, Eastern Province [9,44] Burundi: Near Lake Shohoho and Rugweru region [44] |
| Southern Africa | Namibia, Zambia, Zimbabwe, Mozambique, South Africa, Malawi, Swaziland, Botswana | South Africa: Eastern Cape; Free state, Gauteng, Kwazulu-Natal [7,44,52] |
| Central Africa | Chad, Lesotho | Not reported in literature |
| North Africa | Algeria, Ethiopia, Somalia, Morocco, Tunisia, Gibraltar | Ethiopia: Shoa, Domak, Efat, Roth, Adua, Mt. Sholoda, Tigray [30] Somalia: Surud, Mt. Maydh, Mt. Hildebrandt [30] Algeria: Tangiers [8,44] |
| Cross-border Islands | Socotra | Socotra: Yemen, Haghier Hills [44] |
| Use Category | Plant Resource | Description |
|---|---|---|
| Cultural/spiritual uses/perfumery | Wood and oil | Oils are used to make perfumes and cosmetics [112] while the wood is burnt during ceremonies in Asia [4] |
| Pharmaceutical/local medicine | Leaves, bark, root | Oils are used in pharmaceutical industries [112], local decoctions to treat malaria [53] and for sexually transmitted diseases (STDs) [32], chest pain, hepatitis B, fever, diarrhoea, chronic mucus infections, cough and asthma [113] |
| Tanning and dyeing | Roots | Roots contain a red dye used for tanning leather in Africa [114] |
| Food | Root, bark oil extract; fruits | Roots and bark provide flavored powder for tea and are used as tonics The root extract is used in preservation of milk in Africa, while the fruits are eaten as emergency foodstuffs by children and herdsmen in east Africa [113,114,115] |
| Ecological services | Root system | Root haustoria can accumulate heavy metals for phytoremediation [1,116] |
| Timber | Wood | Hardwood is used to make carvings and fencing for homesteads in Africa [5,8] |
| Ethnoveterinary uses | Leaves | The leaves are used as fodder, and also contain antipyretic agents for cattle in east Africa [117] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mugula, B.B.; Kiboi, S.K.; Kanya, J.I.; Egeru, A.; Okullo, P.; Curto, M.; Meimberg, H. Knowledge Gaps in Taxonomy, Ecology, Population Distribution Drivers and Genetic Diversity of African Sandalwood (Osyris lanceolata Hochst. & Steud.): A Scoping Review for Conservation. Plants 2021, 10, 1780. https://doi.org/10.3390/plants10091780
Mugula BB, Kiboi SK, Kanya JI, Egeru A, Okullo P, Curto M, Meimberg H. Knowledge Gaps in Taxonomy, Ecology, Population Distribution Drivers and Genetic Diversity of African Sandalwood (Osyris lanceolata Hochst. & Steud.): A Scoping Review for Conservation. Plants. 2021; 10(9):1780. https://doi.org/10.3390/plants10091780
Chicago/Turabian StyleMugula, Ben Belden, Samuel Kuria Kiboi, James Ireri Kanya, Anthony Egeru, Paul Okullo, Manuel Curto, and Harald Meimberg. 2021. "Knowledge Gaps in Taxonomy, Ecology, Population Distribution Drivers and Genetic Diversity of African Sandalwood (Osyris lanceolata Hochst. & Steud.): A Scoping Review for Conservation" Plants 10, no. 9: 1780. https://doi.org/10.3390/plants10091780