
Rice Phytoalexins: Half a Century of Amazing Discoveries; Part I: Distribution, Biosynthesis, Chemical Synthesis, and Biological Activities


Abstract
:
1. Introduction
2. Diversity and Distribution of Rice Phytoalexins
2.1. Diterpenoid Phytoalexins in Rice
2.1.1. Momilactones
2.1.2. Phytocassanes
2.1.3. Oryzalexins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Species (Subspecies, Cultivar) | Part of the Plant | References |
|---|---|---|---|
| Momilactones A and B | Oryza sativa L. (subsp. japonica cv. Koshihikari) | Seed husks | [27,55] |
| O. sativa (subsp. japonica cvs. Sasashigure; Koshihikari; Haresugata) | Leaves and straw | [28,54,56,57] | |
| O. sativa (subsp. japonica cv. Koshihikari) | Root exudates | [58,59,60,61] | |
| Wild rice species: Oryza barthii A. Chev.; O. brachyantha A. Chev. et Rhoer.; O. glaberrima Steud.; O. glumaepatula Steud.; O. meridionalis N. Q. Ng; O. punctata Kotschy ex Steud.; O. rufipogon Griff. | UV-light-irradiated leaves | [9,44] | |
| Echinochloa crus-galli (L.) P.Beauv. | Leaves | [45,46] | |
| Calohypnum plumiforme (Wilson) Jan Kučera & Ignatov (formerly Hypnum plumaeforme Wilson) | Aerial parts | [48,62,63,64,65] | |
| Plagiomnium acutum (Lindb.) T. Kop. | Plant material | [47] | |
| Momilactone C | O. sativa (subsp. japonica cv. Koshihikari) | Seed husks | [42] |
| Pseudoleskeella papillosa (Lindb.) Kindb | Plant material | [66] | |
| Momilactones D and E | O. sativa (subsp. japonica cv. Chucheongbyeo) | Roots | [43] |
| Oryzalactone | O. sativa (subsp. tropical japonica cv. Jaguary; subsp. indica cvs. Local Basmati and Bingala) Wild rice species: O. rufipogon; O. meridionalis | Leaves irradiated with UV light or inoculated with conidia of Magnaporthe grisea | [9] |
| Phytocassanes A, B, C and D | O. sativa (subsp. japonica cv. Jukkoku) | Leaves infected with Magnaporthe grisea and stems infected with Rhizoctonia solani | [29] |
| Phytocassanes A and D | Wild rice species: Oryza barthii A. Chev.; O. brachyantha A. Chev. et Rhoer.; O. glaberrima Steud.; O. glumaepatula Steud.; O. meridionalis N. Q. Ng; O. rufipogon Griff. | Leaves | [9] |
| Phytocassane E | O. sativa (subsp. japonica cv. Koshihikari) | Suspension-cultured rice cells elicited with Phytophthora infestans mycelial extract | [30] |
| Phytocassane F | O. sativa (subsp. japonica cv. Koshihikari) | UV-irradiated leaves | [31] |
| Phytocassane G | O. sativa (almost all cultivars in the WRC) Wild rice species: Oryza rufipogon; O. glaberrima; O. barthii; O. glumaepatula; O. meridionalis | UV-irradiated leaves | [9] |
| Oryzalexin A | O. sativa (−) | Leaves infected with Magnaporthe grisea (Syn. Pyricularia oryzae) | [32] |
| O. sativa (69 cultivars in the world rice core collection) and O. rufipogon (10 strains within the clades, Or-I, Or-II, or Or-III) | Leaves irradiated by UV light, treated with jasmonic acid, or inoculated with conidia of Bipolaris oryzae | [53] | |
| Oryzalexins B and C | O. sativa (subsp. japonica cv. Koshihikari) | Leaves infected with M. grisea | [33,34] |
| Oryzalexins D | O. sativa (subsp. japonica cv. Koganenishiki) | Leaves infected with M. grisea | [36] |
| Oryzalexin E | O. sativa (subsp. japonica cv. Nipponbare) | UV-irradiated leaves | [37] |
| Oryzalexin F | O. sativa (subsp. japonica cv. Nipponbare) | UV-irradiated leaves | [38] |
| Oryzalexin S | O. sativa (subsp. japonica cv. Koshihikari) | UV-irradiated leaves | [39,40,54] |
2.2. Phenolic Phytoalexins in Rice
| Species | Family | Part of the Plant | Reference |
|---|---|---|---|
| Prunus pseudocerasus Lindl. | Rosaceae | Bark | [68] |
| Prunus cerasoides D.Don (Prunus puddum) | Rosaceae | Bark | [69] |
| Eupatorium havanense Kunth | Asteraceae | Whole plant | [75] |
| Ribes nigrum L. | Grossulariaceae | Leaves | [76] |
| Iris milesii Baker ex Foster | Iridaceae | Rhizomes | [77] |
| Artemisia campestris subsp. glutinosa (Gay ex Bess.) Batt. | Asteraceae | Aerial parts | [78] |
| Hyptis salzmanii Benth. | Lamiaceae | Leaves | [79] |
| Bonnetia dinizii Huber | Guttiferae | Wood | [80] |
| Primula sieboldii E. Morren | Primulaceae | Bud exudate | [81] |
| Eriodictyon californicum (Hook. & Arn.) Torr. | Boraginaceae | Leaves | [82] |
| Teucrium stocksianum Boiss. | Lamiaceae | Aerial parts | [83] |
| Dodonaea viscosa Jacq. | Sapindaceae | Aerial parts | [84] |
| Xanthorrhoea hastilis R. Br. | Xanthorrhoeaceae | Dried resin | [85] |
| Daphne aurantiaca Diels. | Thymelaeaceae | Stem bark | [86] |
| Dodonaea viscosa Jacq. | Sapindaceae | Aerial parts | [87] |
| Baccharis retusa DC. | Asteraceae | Twigs | [88] |
| Dicerothamnus rhinocerotis Less. | Asteraceae | Leaves | [89] |
| Prunus avium L. | Rosaceae | Sweet cherry | [90] |
| Viscum album L. | Santalaceae | Tinctures | [91] |
3. Rice Phytoalexin Biosynthesis
3.1. Biosynthesis of Rice Diterpenoid Phytoalexins


3.1.1. Biosynthesis of Momilactones
3.1.2. Biosynthesis of Phytocassanes
3.1.3. Biosynthesis of Oryzalexins
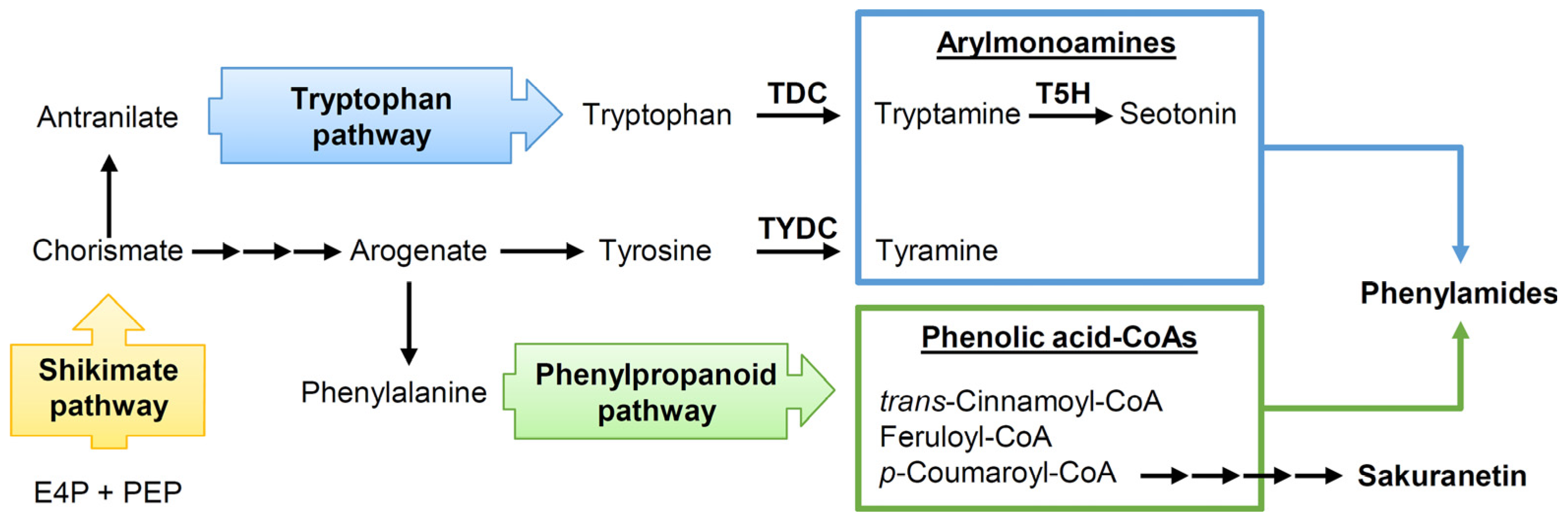
3.2. Biosynthesis of Rice Flavonoid Phytoalexins
4. Chemical Synthesis of Rice Phytoalexins
4.1. Momilactones Chemical Synthesis
4.2. Phytocassane Chemical Synthesis
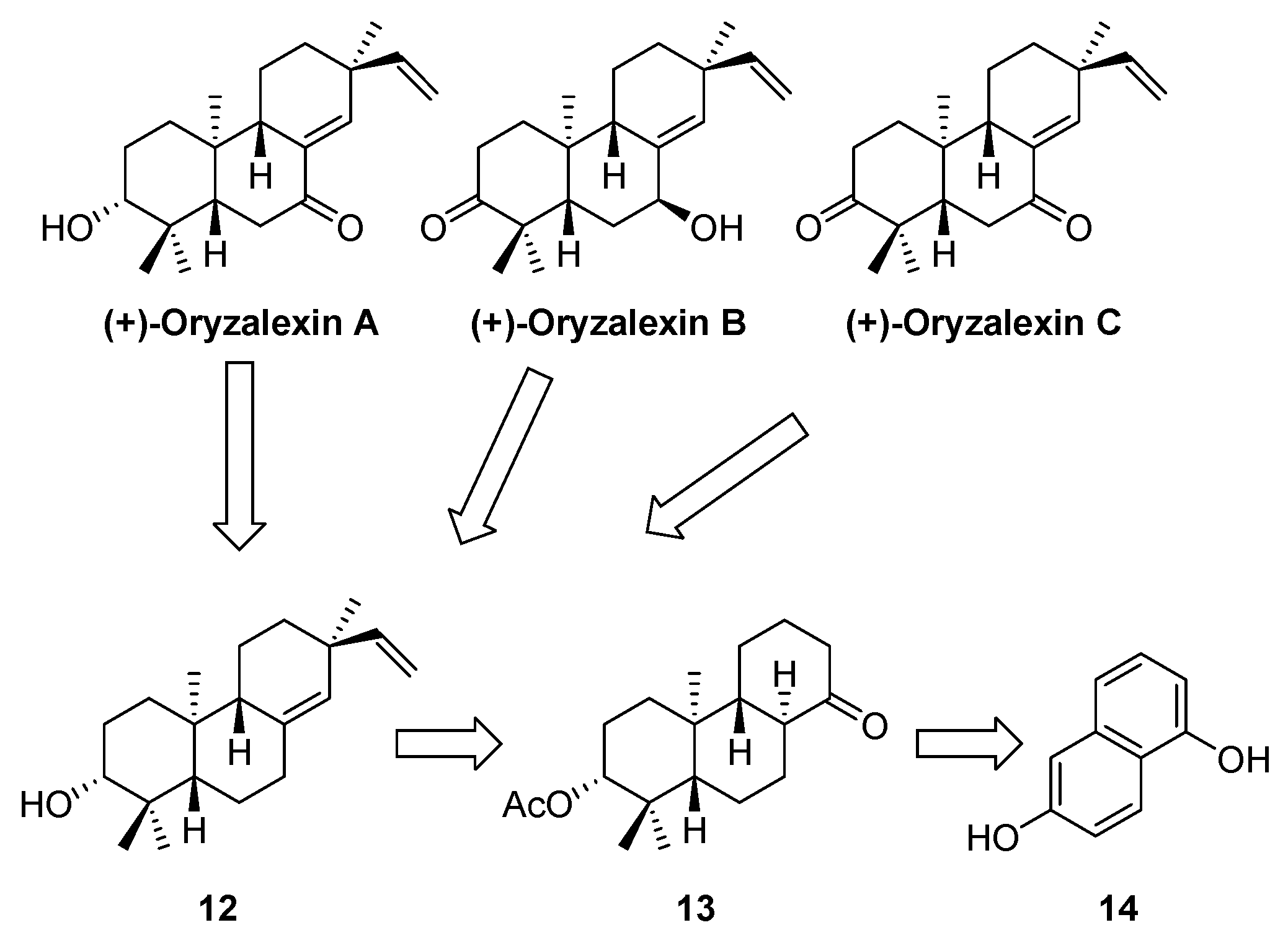
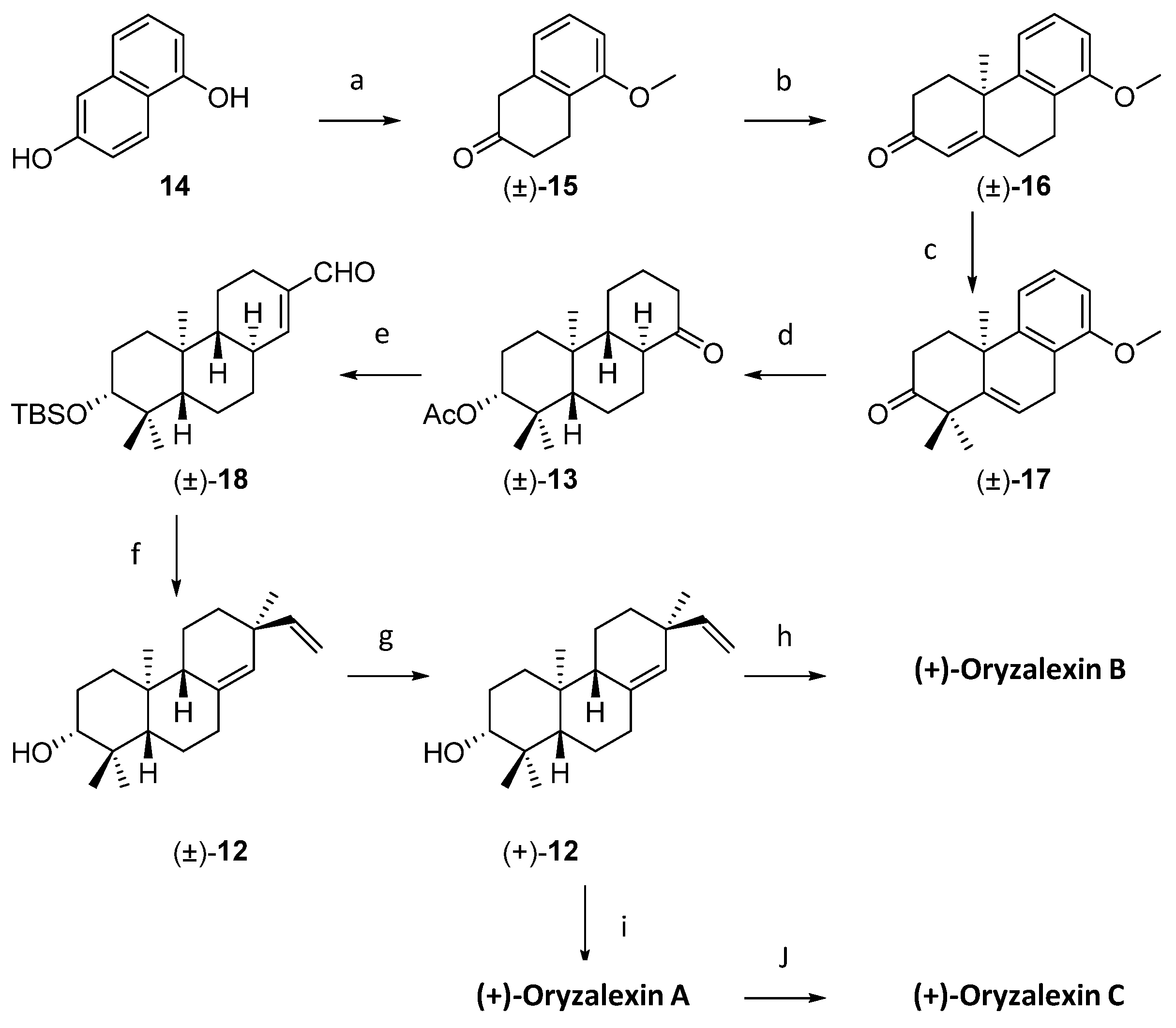
4.3. Oryzalexin Chemical Synthesis
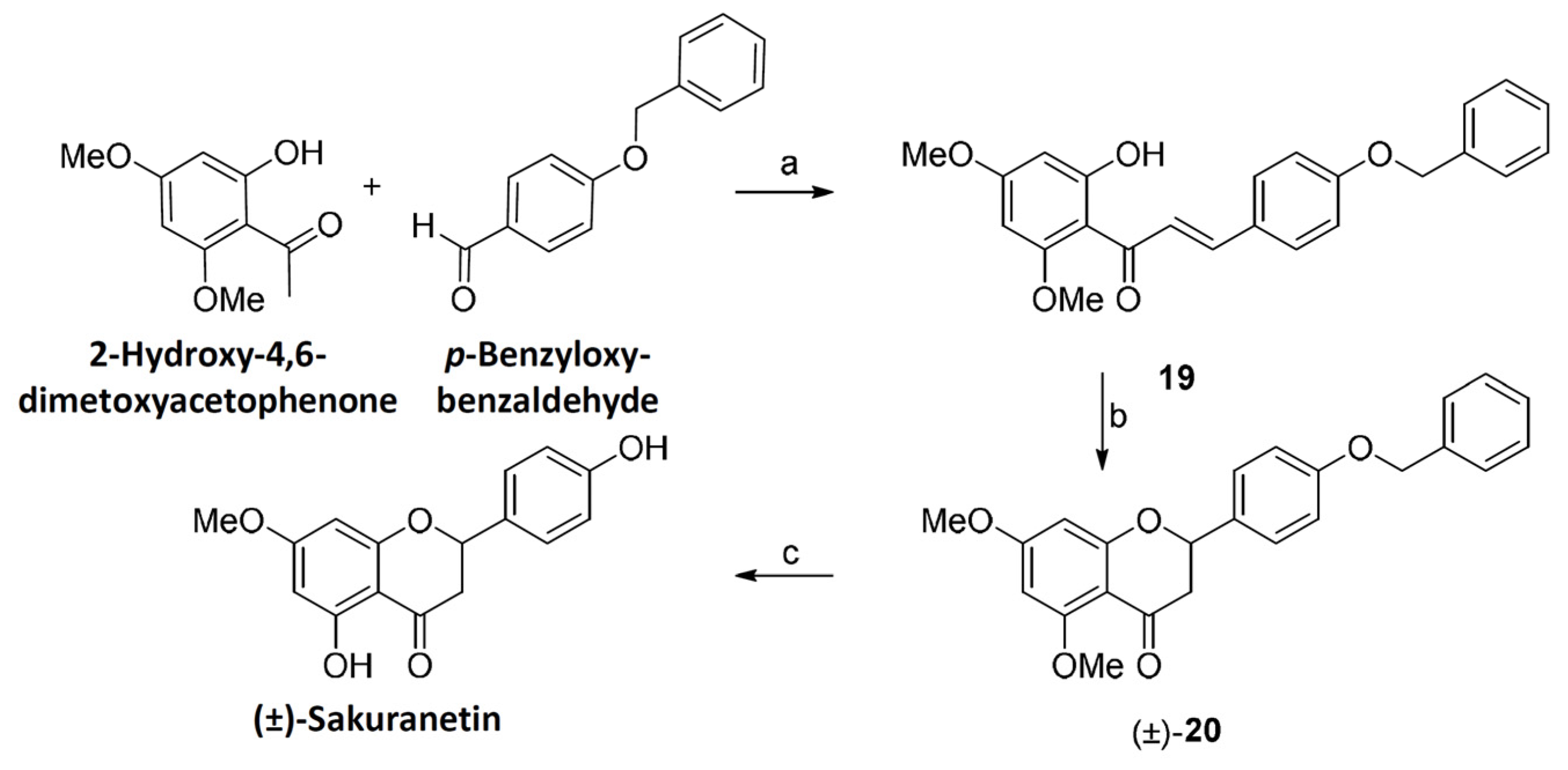
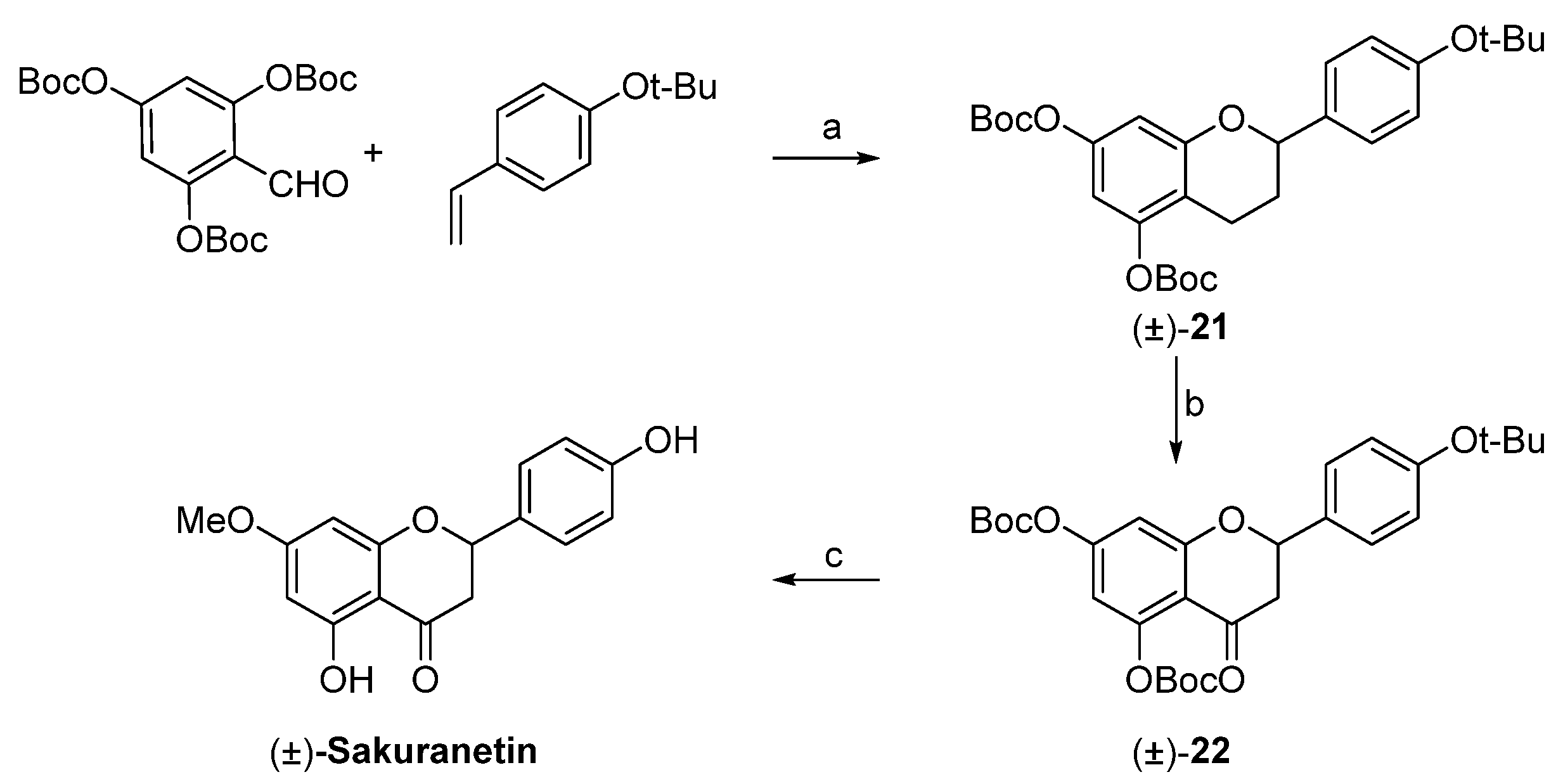
4.4. Sakuranetin Chemical Synthesis
5. Biological Activities of Rice Phytoalexins
5.1. Bioactivities of Diterpenoid Phytoalexins
5.1.1. Momilactones
- Antimicrobial and allelopathic activity:
- Anti-cancer activity:
- Other activities:
5.1.2. Oryzalexins
5.1.3. Phytocassanes
5.2. Bioactivities of Phenolic Phytoalexins
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3LN0 | Structure of compound 5c-S bound at the active site of COX-2 |
| 4HZ5 | Pyrrolopyrimidine inhibitors of DNA gyrase B and topoisomerase iv |
| 4KIK | Human IkB kinase beta |
| 5-LOX | 5-Lipoxygenase |
| ACC | Acetyl-CoA carboxylase |
| ADP | Adenosine diphosphate |
| AF-2 | Furylfuramide |
| BenSer | N-Benzoylserotonin |
| BenTry | N-Benzoyltryptamine |
| BenTyr | N-Benzoyltyramine |
| BGC | Biosynthetic gene cluster |
| CDP | Copalyl diphosphate |
| CHI | Chalcone isomerase |
| CHS | Chalcone synthase |
| CinSer | N-trans-Cinnamoylserotonin |
| Cin-Try | N-trans-Cinnamoyltryptamine |
| Cin-Tyr | N-trans-Cinnamoyltyramine |
| CM | Chorismate mutase |
| CouSer | p-Coumaroylserotonin |
| CS | Chorismate synthase |
| cv. | Cultivar |
| CYPs | Cytochrome P450 enzymes |
| DAHP | 3-Deoxy-D-arabinoheptulosonate 7-phosphate |
| DAHPS | 3-Deoxy-D-arabinoheptulosonate 7-phosphate synthase |
| DHQDT/SDH | 3-Dehydroquinate dehydratase/shikimate dehydrogenase |
| DHQS | 3-Dehydroquinate synthase |
| diTPS | Diterpene-synthase |
| DMAPP | Dimethylallyl diphosphate |
| EC50 | Half maximal effective concentration |
| ent-CPP | (5R,9S,10S)-Copalyl diphosphate |
| ER | Endoplasmic reticulum |
| FerAgm | N-Feruloylagmatine |
| FerPut | N-Feruloylputrescine |
| FerSer | N-Feruloylserotonin |
| FerTry | N-Feruloyltryptamine |
| FPP | Farnesyl diphosphate |
| GAs | Gibberellins |
| GFP | Green fluorescent protein |
| GGPP | Geranylgeranyl diphosphate |
| GGPPS | Geranylgeranyl diphosphate synthases |
| GPP | Geranyl diphosphate |
| HDR | 4-Hydroxy-3-methylbut-2-enyl diphosphate reductase |
| HEL | Hen egg-white lysozyme |
| HMBPP | 4-Hydroxy-3-methylbut-2-enyl diphosphate |
| IC50 | Half maximal inhibitory concentration |
| IDS | Isoprenyl diphosphate synthase |
| IPP | Isopentenyl diphosphate |
| IPPI | Isopentenyl diphosphate isomerase |
| IRRI | International Rice Research Institute |
| JA | Jasmonic acid |
| LPS | Lipopolysaccharides |
| LTB4 | Leukotriene B4 |
| MEP | Methylerythritol 4-phosphate |
| MPO | Myeloperoxidase |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-419 2,5-diphenyltetrazolium bromide |
| MVA | Mevalonate |
| NO | Nitric oxide |
| NOMT | Naringenin 7-O-methyltransferase |
| OMT | Flavonoid O-methyltransferase |
| Phe | Phenylalanine |
| PKC | Protein kinase C |
| PLA2 | Phospholipase A2 |
| PPi | Pyrophosphate |
| PRs | Pathogenesis-related proteins |
| ROS | Reactive oxygen species |
| SAH | Adenosyl-L-homocysteine |
| SAM | S-Adenosyl-L-methionine |
| SDR | Short-chain dehydrogenase reductase |
| SK | Shikimate kinase |
| syn-CPP | (5S,9S,10R)-Copalyl diphosphate |
| TDC | Tryptophan decarboxylase |
| TPA | 12-O-Tetradecanoylphorbol 13-acetate |
| Try | Tryptophan |
| TYDC | Tyrosine decarboxylase |
| Tyr | Tyrosine |
| UBQ5 | Ubiquinone 5 |
| UV | Ultraviolet radiation |
| VEGF | Vascular endothelial growth factor |
| WBA | Whole blood aggregometer |
| WRC | World rice core collection |
References
- Wang, W.; Li, Y.; Dang, P.; Zhao, S.; Lai, D.; Zhou, L. Rice secondary metabolites: Structures, roles, biosynthesis, and metabolic regulation. Molecules 2018, 23, 3098. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.Y.; Liu, X.L.; Xiao, Y.H.; Wang, G.L. Recent advances in cloning and characterization of disease resistance genes in rice. J. Integr. Plant Biol. 2007, 49, 112–119. [Google Scholar] [CrossRef]
- Sparks, A.; Nelson, A.; Castilla, N. Where rice pests and diseases do the most damage. Rice Today 2012, 11, 27. [Google Scholar]
- Sun, X.D.; Su, P.; Shan, H. Mycotoxin contamination of rice in China. J. Food Sci. 2017, 82, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Müller, K.O.; Meyer, G.; Klinkowski, M. Physiologisch-genetische Untersuchungen über die Resistenz der Kartoffel gegenüber Phytophthora infestans. Naturwissenschaften 1939, 27, 765–768. [Google Scholar] [CrossRef]
- Müller, K.O.; Börger, H. Experimentelle Untersuchungen über die Phytophthora-Resistenz der Kartoffel. Arbeit. Biol. Reichsant Land Forstwirtsch. 1940, 23, 189–231. [Google Scholar]
- Stoessl, A. Phytoalexins—A Biogenetic Perspective. Phytopathol. Z. 1980, 99, 251–272. [Google Scholar] [CrossRef]
- VanEtten, H.D.; Mansfield, J.W.; Bailey, J.A.; Farmer, E.E. Two classes of plant antibiotics: Phytoalexins versus “phytoanticipins”. Plant Cell 1994, 6, 1191. [Google Scholar] [CrossRef]
- Kariya, K.; Ube, N.; Ueno, M.; Teraishi, M.; Okumoto, Y.; Mori, N.; Ueno, K.; Ishihara, A. Natural variation of diterpenoid phytoalexins in cultivated and wild rice species. Phytochemistry 2020, 180, 112518. [Google Scholar] [CrossRef]
- Estiati, A. Rice momilactones, potential allelochemical for weeds suppression. Asian J. Agric. 2019, 3, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Pang, Y.; Lu, F.; Ding, C.; Zeng, R.; Song, Y. Review on momilactones of key allelochemicals in rice allelopathy. Guangxi Zhiwu/Guihaia 2019, 39, 548–556. [Google Scholar]
- Serra Serra, N.; Shanmuganathan, R.; Becker, C. Allelopathy in rice: A story of momilactones, kin recognition, and weed management. J. Exp. Bot. 2021, 72, 4022–4037. [Google Scholar] [CrossRef] [PubMed]
- Aci, M.M.; Sidari, R.; Araniti, F.; Lupini, A. Emerging Trends in Allelopathy: A Genetic Perspective for Sustainable Agriculture. Agronomy 2022, 12, 2043. [Google Scholar] [CrossRef]
- Sindhu, R.K.; Arora, B.; Arora, S. Phytoalexins: Sources and their pharmacological potential. J. Pharm. Technol. Res. Manag. 2020, 8, 1–8. [Google Scholar] [CrossRef]
- Bizuneh, G.K. The chemical diversity and biological activities of phytoalexins. Adv. Tradit. Med. 2021, 21, 31–43. [Google Scholar] [CrossRef]
- Choi, H.J. In vitro antiviral activity of sakuranetin against human rhinovirus 3. Osong Public Health Res. Perspect. 2017, 8, 415–420. [Google Scholar] [CrossRef]
- Kwon, D.H.; Ji, J.H.; Yim, S.H.; Kim, B.S.; Choi, H.J. Suppression of influenza B virus replication by sakuranetin and mode of its action. Phytother. Res. 2018, 32, 2475–2479. [Google Scholar] [CrossRef]
- Park, J.H.; Fu, Y.Y.; Chung, I.S.; Hahn, T.R.; Cho, M.H. Cytotoxic property of ultraviolet-induced rice phytoalexins to human colon carcinoma HCT-116 cell. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 237–241. [Google Scholar] [CrossRef]
- Fukuta, M.; Xuan, T.D.; Deba, F.; Tawata, S.; Khanh, T.D.; Chung, I.M. Comparative efficacies in vitro of antibacterial, fungicidal, antioxidant, and herbicidal activities of momilatones A and B. J. Plant Interact. 2007, 2, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Cheng, J.; Guo, B.; Duan, J.; Che, C.T. Momilactone and related diterpenoids as potential agricultural chemicals. J. Agric. Food Chem. 2018, 66, 7859–7872. [Google Scholar] [CrossRef]
- Cho, M.H.; Lee, S.W. Phenolic phytoalexins in rice: Biological functions and biosynthesis. Int. J. Mol. Sci. 2015, 16, 29120–29133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, K.M.; Zerbe, P. Specialized diterpenoid metabolism in monocot crops: Biosynthesis and chemical diversity. Phytochemistry 2020, 172, 112289. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Liu, X.; Tian, M.; Ma, Y.; Jin, B.; Gao, W.; Cui, G.; Guo, J.; Huang, L. Recent progress and new perspectives for diterpenoid biosynthesis in medicinal plants. Med. Res. Rev. 2021, 41, 2971–2997. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.J. Two rings in them all: The labdane-related diterpenoids. Nat. Prod. Rep. 2010, 27, 1521–1530. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Zha, W.L.; Zhu, J.X.; Zheng, C.; Zi, J.C. A review: Biosynthesis of plant-derived labdane-related diterpenoids. Chin. J. Nat. Med. 2021, 19, 666–674. [Google Scholar] [CrossRef]
- Murphy, K.M.; Ma, L.T.; Ding, Y.; Schmelz, E.A.; Zerbe, P. Functional characterization of two class II diterpene synthases indicates additional specialized diterpenoid pathways in maize (Zea mays). Front. Plant Sci. 2018, 9, 1542. [Google Scholar] [CrossRef] [Green Version]
- Kato, T.; Kabuto, C.; Sasaki, N.; Tsunagawa, M.; Aizawa, H.; Fujita, K.; Kato, Y.; Kitahara, Y.; Takahashi, N. Momilactones, growth inhibitors from rice, Oryza sativa L. Tetrahedron Lett. 1973, 14, 3861–3864. [Google Scholar] [CrossRef]
- Cartwright, D.W.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Isolation and characterization of two phytoalexins from rice as momilactones A and B. Phytochemistry 1981, 20, 535–537. [Google Scholar] [CrossRef]
- Koga, J.; Shimura, M.; Oshima, K.; Ogawa, N.; Yamauchi, T.; Ogasawara, N. Phytocassanes A, B, C and D, novel diterpene phytoalexins from rice, Oryza sativa L. Tetrahedron 1995, 51, 7907–7918. [Google Scholar] [CrossRef]
- Koga, J.; Ogawa, N.; Yamauchi, T.; Kikuchi, M.; Ogasawara, N.; Shimura, M. Functional moiety for the antifungal activity of phytocassane E, a diterpene phytoalexin from rice. Phytochemistry 1997, 44, 249–253. [Google Scholar] [CrossRef]
- Horie, K.; Inoue, Y.; Sakai, M.; Yao, Q.; Tanimoto, Y.; Koga, J.; Toshima, H.; Hasegawa, M. Identification of UV-induced diterpenes including a new diterpene phytoalexin, phytocassane F, from rice leaves by complementary GC/MS and LC/MS approaches. J. Agric. Food Chem. 2015, 63, 4050–4059. [Google Scholar] [CrossRef] [PubMed]
- Akatsuka, T.; Kodama, O.; Kato, H.; Kono, Y.; Takeuchi, S. 3-Hydroxy-7-oxo-sandaracopimaradiene (oryzalexin A), a new phytoalexin isolated from rice blast leaves. Agric. Biol. Chem. 1983, 47, 445–447. [Google Scholar] [CrossRef] [Green Version]
- Akatsuka, T.; Kodama, O.; Sekido, H.; Kono, Y.; Takeuchi, S. Novel phytoalexins (oryzalexins A, B and C) isolated from rice blast leaves infected with Pyricularia oryzae. Part I: Isolation, characterization and biological activities of oryzalexins. Agric. Biol. Chem. 1985, 49, 1689–1694. [Google Scholar] [CrossRef]
- Kono, Y.; Takeuchi, S.; Kodama, O.; Akatsuka, T. Absolute configuration of oryzalexin A and structures of its related phytoalexins isolated from rice blast leaves infected with Pyricularia oryzae. Agric. Biol. Chem. 1984, 48, 253–255. [Google Scholar] [CrossRef]
- Kono, Y.; Takeuchi, S.; Kodama, O.; Sekido, H.; Akatsuka, T. Novel phytoalexins (oryzalexins A, B and C) isolated from rice blast leaves infected with Pyricularia oryzae. Part II: Structural studies of oryzalexins. Agric. Biol. Chem. 1985, 49, 1695–1701. [Google Scholar] [CrossRef]
- Sekido, H.; Endo, T.; Suga, R.; Kodama, O.; Akatsuka, T.; Kono, Y.; Takeuchi, S. Oryzalexin D (3, 7-dihydroxy-(+)-sandaracopimaradiene), a new phytoalexin isolated from blast-infected rice leaves. J. Pestic. Sci. 1986, 11, 369–372. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Kodama, O.; Akatsuka, T. Oryzalexin E, a diterpene phytoalexin from UV-irradiated rice leaves. Phytochemistry 1993, 33, 79–81. [Google Scholar] [CrossRef]
- Kato, H.; Kodama, O.; Akatsuka, T. Oryzalexin F, a diterpene phytoalexin from UV-irradiated rice leaves. Phytochemistry 1994, 36, 299–301. [Google Scholar] [CrossRef]
- Kodama, O.; Li, W.X.; Tamogami, S.; Akatsuka, T. Oryzalexin S, a novel stemarane-type diterpene rice phytoalexin. Biosci. Biotechnol. Biochem. 1992, 56, 1002–1003. [Google Scholar] [CrossRef]
- Tamogami, S.; Mitani, M.; Kodama, O.; Akatsuka, T. Oryzalexin S structure: A new stemarane-type rice plant phytoalexin and its biogenesis. Tetrahedron 1993, 49, 2025–2032. [Google Scholar] [CrossRef]
- Gu, C.Z.; Xia, X.M.; Lv, J.; Tan, J.W.; Baerson, S.R.; Pan, Z.Q.; Song, Y.-Y.; Zeng, R.S. Diterpenoids with herbicidal and antifungal activities from hulls of rice (Oryza sativa). Fitoterapia 2019, 136, 104183. [Google Scholar] [CrossRef] [PubMed]
- Tsunakawa, M.; Ohba, A.; Sasaki, N.; Kabuto, C.; Kato, T.; Kitahara, Y.; Takahashi, N. Momilactone C, a minor constituent of growth inhibitors in rice husk. Chem. Lett. 1976, 5, 1157–1158. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.G.; Cha, B.J.; Lee, S.M.; Shrestha, S.; Jeong, R.H.; Lee, D.S.; Kim, Y.C.; Lee, D.G.; Kang, H.C.; Kim, J.; et al. Diterpenes from the roots of Oryza sativa L. and their inhibition activity on NO production in LPS-stimulated RAW264.7 macrophages. Chem. Biodiversity 2015, 12, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Fujita, M.; Shenton, M.F.; Shenton, M.R.; Akashi, S.; Sugawara, C.; Sakai, A.; Horie, K.; Hasegawa, M.; Kawide, H.; et al. Evolutionary trajectory of phytoalexin biosynthetic gene clusters in rice. Plant J. 2016, 87, 293–304. [Google Scholar] [CrossRef]
- Guo, L.; Qiu, J.; Ye, C.; Jin, G.; Mao, L.; Zhang, H.; Yang, X.; Peng, Q.; Wang, Y.; Longjiang, F.; et al. Echinochloa crus-galli genome analysis provides insight into its adaptation and invasiveness as a weed. Nat. Commun. 2017, 8, 1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Peters, R.J. Why are momilactones always associated with biosynthetic gene clusters in plants? Proc. Natl. Acad. Sci. USA 2020, 117, 13867–13869. [Google Scholar] [CrossRef]
- Toyota, M.; Kimura, K.; Asakawa, Y. Occurence of ent-sesquiterpene in the japanese moss Plagiomnium acutum: First isolation and identification of the ent-sesqui- and dolabellane-type diterpenoids from the musci. Chem. Pharm. Bull. 1998, 46, 1488–1489. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, H.; Hayashi, K.I.; Nishimura, N.; Kawaide, H.; Matsuo, A.; Takaoka, D. Momilactone A and B as allelochemicals from moss Hypnum plumaeforme: First occurrence in bryophytes. Biosci. Biotechnol. Biochem. 2007, 71, 3127–3130. [Google Scholar] [CrossRef]
- Kojima, Y.; Ebana, K.; Fukuoka, S.; Nagamine, T.; Kawase, M. Development of an RFLP-based rice diversity research set of germplasm. Breed. Sci. 2005, 55, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Toyomasu, T.; Kagahara, T.; Okada, K.; Koga, J.; Hasegawa, M.; Mitsuhashi, W.; Sassa, T.; Yamane, H. Diterpene phytoalexins are biosynthesized in and exuded from the roots of rice seedlings. Biosci. Biotechnol. Biochem. 2008, 72, 562–567. [Google Scholar] [CrossRef]
- Sesma, A.; Osbourn, A.E. The rice leaf blast pathogen undergoes developmental processes typical of root-infecting fungi. Nature 2004, 431, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Khush, G.S. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol. 1997, 35, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Kariya, K.; Murata, K.; Kokubo, Y.; Ube, N.; Ueno, K.; Yabuta, Y.; Teraishi, M.; Okumoto, Y.; Mori, N.; Ishihara, A. Variation of diterpenoid phytoalexin oryzalexin A production in cultivated and wild rice. Phytochemistry 2019, 166, 112057. [Google Scholar] [CrossRef] [PubMed]
- Kodama, O.; Yamada, A.; Yamamoto, A.; Takemoto, T.; Alkatsuka, T. Induction of Phytoalexins with Heavy Metal Ions in Rice Leaves. J. Pestic. Sci. 1988, 13, 615–617. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Kato, T.; Tsunagawa, M.; Sasaki, N.; Kitahara, Y. Mechanisms of dormancy in rice seeds: II. New growth inhibitors, momilactone-A and-B isolated from the hulls of rice seeds. Jpn. J. Breed. 1976, 26, 91–98. [Google Scholar] [CrossRef]
- Cartwright, D.; Langcake, P.; Pryce, R.J.; Leworthy, D.P.; Ride, J.P. Chemical activation of host defence mechanisms as a basis for crop protection. Nature 1977, 267, 511–513. [Google Scholar] [CrossRef]
- Lee, C.W.; Yoneyama, K.; Takeuchi, Y.; Konnai, M.; Tamogami, S.; Kodama, O. Momilactones A and B in rice straw harvested at different growth stages. Biosci. Biotechnol. Biochem. 1999, 63, 1318–1320. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T.; Sata, N.; Yamamura, S. Isolation and identification of a potent allelopathic substance in rice root exudates. Physiol. Plant. 2002, 115, 401–405. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathic substance in rice root exudates: Rediscovery of momilactone B as an allelochemical. J. Plant Physiol. 2004, 161, 271–276. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T.; Ota, K. Secretion of momilactone A from rice roots to the rhizosphere. J. Plant Physiol. 2008, 165, 691–696. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ota, K.; Ino, T. Release of momilactone A and B from rice plants into the rhizosphere and its bioactivities. Allelopathy J. 2008, 22, 321–328. [Google Scholar]
- Kobayashi, K.; Shigemori, H.; Kato-Noguchi, H. Allelopathic potential of Hypnum plumaeforme L. and its allelopathic substances. In Proceedings of the 4th Asia-Pacific Conference on Chemical Ecology, from Biomolecules to Ecosystems an Interactive Chemical Message for our Future, Tsukuba, Japan, 10–14 September 2007; p. 77. [Google Scholar]
- Kato-Noguchi, H. Secretion of momilactone A and B by the moss Hypnum plumaeforme. Plant Signaling Behav. 2009, 4, 737–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato-Noguchi, H.; Kobayashi, K. Jasmonic acid, protein phosphatase inhibitor, metals and UV-irradiation increased momilactone A and B concentrations in the moss Hypnum plumaeforme. J. Plant Physiol. 2009, 166, 1118–1122. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kobayashi, K.; Shigemori, H. Allelopathy of the moss Hypnum plumaeforme by the production of momilactone A and B. Weed Res. 2009, 49, 621–627. [Google Scholar] [CrossRef]
- Liu, N.; Wang, S.; Lou, H. A new pimarane-type diterpenoid from moss Pseudoleskeella papillosa (Lindb.) Kindb. Acta Pharm. Sin. B. 2012, 2, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Venkateswara Rao, P.; Kiran, S.; Rohini, P.; Bhagyasree, P. Flavonoid: A review on Naringenin. J. Pharmacogn. Phytochem. 2017, 6, 2778–2783. [Google Scholar]
- Asahina, Y. Ueber das sakuranin, ein neues glykosid der rinde von Prunus pseudo-Cerasus lindl. var. sieboldi Maxim. Arch. Pharm. 1908, 246, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Narasimhachabi, N.; Seshadbi, T.R. A note on the components of the bark of Prunus puddum. Proc. Indian Acad. Sci. A 1949, 30, 274–276. [Google Scholar]
- Kodama, O.; Miyakawa, J.; Akatsuka, T.; Kiyosawa, S. Sakuranetin, a flavanone phytoalexin from ultraviolet-irradiated rice leaves. Phytochemistry 1992, 31, 3807–3809. [Google Scholar] [CrossRef]
- Stompor, M. A review on sources and pharmacological aspects of sakuranetin. Nutrients 2020, 12, 513. [Google Scholar] [CrossRef] [Green Version]
- Dillon, V.M.; Overton, J.; Grayer, R.J.; Harborne, J.B. Differences in phytoalexin response among rice cultivars of different resistance to blast. Phytochemistry 1997, 44, 599–603. [Google Scholar] [CrossRef]
- Hasegawa, M.; Mitsuhara, I.; Seo, S.; Okada, K.; Yamane, H.; Iwai, T.; Ohashi, Y. Analysis on blast fungus-responsive characters of a flavonoid phytoalexin sakuranetin; accumulation in infected rice leaves, antifungal activity and detoxification by fungus. Molecules 2014, 19, 11404–11418. [Google Scholar] [CrossRef] [PubMed]
- Park, H.L.; Yoo, Y.; Hahn, T.R.; Bhoo, S.H.; Lee, S.W.; Cho, M.H. Antimicrobial activity of UV-induced phenylamides from rice leaves. Molecules 2014, 19, 18139–18151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez, X.A.; De La Fuente, E.R. Sakuranetin and pulcherryl acetate from Eupatorium havanense. Phytochemistry 1973, 12, 2060. [Google Scholar] [CrossRef]
- Atkinson, P.; Blakeman, J.P. Seasonal occurrence of an antimicrobial flavanone, sakuranetin, associated with glands on leaves of Ribes nigrum. New Phytol. 1982, 92, 63–74. [Google Scholar] [CrossRef]
- Agrwal, V.K.; Thappa, R.K.; Agrwal, S.G.; Ghar, K.L. Phenolic constituents of Iris milessi rhizomes. Phytochemistry 1984, 23, 1342–1343. [Google Scholar] [CrossRef]
- De Pascual, T.; González, M.S.; Muriel, M.R.; Bellido, I.S. Phenolic derivatives from Artemisia campestris subsp. glutinosa. Phytochemistry 1984, 23, 1819–1821. [Google Scholar] [CrossRef]
- Messana, I.; Ferrari, F.; de Moraes e Souza, M.A.; Gács-Baitz, E. (-)-Salzol, an isopimarane diterpene, and a chalcone from Hyptis salzmanii. Phytochemistry 1990, 29, 329–332. [Google Scholar] [CrossRef]
- De Oliveira, W.G.; Mesquita, A.A.L.; Kubitzki, K.; Gottlieb, O.R. Xanthones from Bonnetia dinizii. Phytochemistry 1990, 29, 1893–1894. [Google Scholar] [CrossRef]
- Greenaway, W.; English, S.; May, J.; Whatley, F.R. Analysis of phenolic of bud exudate of Populus sieboldii by GC-MS. Phytochemistry 1991, 30, 3005–3008. [Google Scholar] [CrossRef]
- Liu, Y.L.; Ho, D.K.; Cassady, J.M.; Cook, V.M.; Baird, W.M. Isolation of potential cancer chemopreventive agents from Eriodictyon californicum. J. Nat. Prod. 1992, 55, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Bashir, A.K. Methylated flavones of Teucrium stocksianum. J. Herbs Spices Med. Plants 1995, 3, 17–24. [Google Scholar] [CrossRef]
- Rojas, A.; Cruz, S.; Ponce-Monter, H.; Mata, R. Smooth muscle relaxing compounds from Dodonaea viscosa. Planta Med. 1996, 62, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Oku, H.; Iwaoka, E.; Inuma, M.; Ishiguro, K. Allergy-preventive flavonoids from Xanthorrhoea hastilis. Chem. Pharm. Bull. 2007, 55, 675–678. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Tian, J.M.; Feng, Y.; Liu, X.H.; Xiong, Z.; Zhang, W.D. Flavonoids from Daphne aurantiaca and their inhibitory activities against nitric oxide production. Chem. Pharm. Bull. 2011, 59, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.B.; Ji, J.; Lei, C.; Wang, H.Y.; Zhao, Q.S.; Hou, A.J. Isoprenylated flavonoid and adipogenesis-promoting constituents of Dodonaea viscosa. J. Nat. Prod. 2012, 75, 699–706. [Google Scholar] [CrossRef]
- Grecco, D.S.; Dorigueto, A.C.; Landre, I.M.; Soares, M.G.; Martho, K.; Lima, R.; Pascon, R.C.; Vallim, M.A.; Capello, T.M.; Romoff, P.; et al. Structural crystalline characterization of sakuranetin—An antimicrobial flavanone from twigs of Baccharis retusa (Asteraceae). Molecules 2014, 19, 7528–7542. [Google Scholar] [CrossRef] [Green Version]
- Ticha, L.A.; Klaasen, J.A.; Green, I.R.; Naidoo, S.; Baker, B.; Pietersen, R.D. Phytochemical and antimicrobial screening of flavanones and chalcones from Galenia africana and Dicerothamnus rhinocerotis. Nat. Prod. Commun. 2015, 10, 1185–1190. [Google Scholar] [CrossRef] [Green Version]
- Aires, A.; Dias, C.; Carvalho, R.; Saavedra, M.J. Analysis of glycosylated flavonoids extracted from sweet-cherry stems, as antibacterial agents against pathogenic Escherichia coli isolates. Acta Biochim. Pol. 2017, 64, 265–271. [Google Scholar] [CrossRef]
- Melo, M.; Oliveira, A.P.; Wiecikowski, A.F.; Castro, J.L.; De Oliveira, A.A.G.; Pereira, H.M.G.; Da Veiga, V.; Capella, M.M.A.; Rocha, L.; Holandino, C. Phenolic compounds from Viscum album tinctures enhanced antitumor activity in melanoma murine cancer cells. Saudi Pharm. J. 2018, 26, 311–322. [Google Scholar] [CrossRef]
- Katsumata, S.; Hamana, K.; Horie, K.; Toshima, H.; Hasegawa, M. Identification of sternbin and naringenin as detoxified metabolites from the rice flavanone phytoalexin sakuranetin by Pyricularia oryzae. Chem. Biodivers. 2017, 14, e1600240. [Google Scholar] [CrossRef] [PubMed]
- Katsumata, S.; Toshima, H.; Hasegawa, M. Xylosylated detoxification of the rice flavonoid phytoalexin sakuranetin by the rice sheath blight fungus Rhizoctonia solani. Molecules 2018, 23, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edreva, A.M.; Velikova, V.B.; Tsonev, T.D. Phenylamides in plants. Russ. J. Plant Physiol. 2007, 54, 287–301. [Google Scholar] [CrossRef]
- Horie, K.; Sakai, K.; Okugi, M.; Toshima, H.; Hasegawa, M. Ultraviolet-induced amides and casbene diterpenoids from rice leaves. Phytochem. Lett. 2016, 15, 57–62. [Google Scholar] [CrossRef]
- Morimoto, N.; Ueno, K.; Teraishi, M.; Okumoto, Y.; Mori, N.; Ishihara, A. Induced phenylamide accumulation in response to pathogen infection and hormone treatment in rice (Oryza sativa). Biosci. Biotechnol. Biochem. 2018, 82, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, A.; Hashmoto, Y.; Tanaka, C.; Dubouzet, J.G.; Nakao, T.; Matsuda, F.; Nishioka, T.; Miyagawa, H.; Wakasa, K. The tryptophan pathway is involved in the defense responses of rice against pathogenic infection via serotonin production. Plant J. 2008, 54, 481–495. [Google Scholar] [CrossRef]
- Ishihara, A.; Nakao, T.; Mashimo, Y.; Murai, M.; Ichimaru, N.; Tanaka, C.; Nakajima, H.; Wakasa, K.; Miyagawa, H. Probing the role of tryptophan-derived secondary metabolism in defense responses against Bipolaris oryzae infection in rice leaves by a suicide substrate of tryptophan decarboxylase. Phytochemistry 2011, 72, 7–13. [Google Scholar] [CrossRef]
- Quinet, M.; Ndayiragije, A.; Lefevre, I.; Lambillotte, B.; Dupont-Gillain, C.C.; Lutts, S. Putrescine differently influences the effect of salt stress on polyamine metabolism and ethylene synthesis in rice cultivars differing in salt resistance. J. Exp. Bot. 2010, 61, 2719–2733. [Google Scholar] [CrossRef]
- Park, H.L.; Lee, S.W.; Jung, K.H.; Hahn, T.R.; Cho, M.H. Transcriptomic analysis of UV-treated rice leaves reveals UV-induced phytoalexin biosynthetic pathways and their regulatory networks in rice. Phytochemistry 2013, 96, 57–71. [Google Scholar] [CrossRef]
- Jin, X.; Baysal, C.; Gao, L.; Medina, V.; Drapal, M.; Ni, X.; Sheng, Y.; Shi, L.; Capell, T.; Fraser, P.D.; et al. The subcellular localization of two isopentenyl diphosphate isomerases in rice suggests a role for the endoplasmic reticulum in isoprenoid biosynthesis. Plant Cell Rep. 2020, 39, 119–133. [Google Scholar] [CrossRef]
- Okada, A.; Shimizu, T.; Okada, K.; Kuzuyama, T.; Koga, J.; Shibuya, N.; Nojiri, H.; Yamane, H. Elicitor induced activation of the methylerythritol phosphate pathway toward phytoalexins biosynthesis in rice. Plant Mol. Biol. 2007, 65, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H. Biosynthesis of phytoalexins and regulatory mechanisms of it in rice. Biosci. Biotechnol. Biochem. 2013, 77, 1141–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemmerlin, A.; Hoeffler, J.F.; Meyer, O.; Tritsch, D.; Kagan, I.A.; Grosdemange-Billiard, C.; Rohmer, M.; Bach, T.J. Cross-talk between the cytosolic mevalonate and the plastidial methyl erythritol phosphate pathways in Tobacco Bright Yellow-2 cells. J. Biol. Chem. 2003, 278, 26666–26676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laule, O.; Fürholz, A.; Chang, H.S.; Zhu, T.; Wang, X.; Heifetz, P.B.; Gruissem, W.; Lange, M. Crosstalk between cytosolic and plastidial pathways of isoprenoid biosynthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6866–6871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-M.; Huang, H.; Chuang, Y.-C.; Chen, W.-H.; Wang, C.-N.; Chen, H.-H. Evolution of Terpene Synthases in Orchidaceae. Int. J. Mol. Sci. 2021, 22, 6947. [Google Scholar] [CrossRef]
- Zerbe, P.; Bohlmann, J. Plant diterpene synthases: Exploring modularity and metabolic diversity for bioengineering. Trends Biotechnol. 2015, 33, 419–428. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Leonelli, F.; Valletta, A.; Migneco, L.M.; Marini Bettolo, R. Stemarane Diterpenes and Diterpenoids. Int. J. Mol. Sci. 2019, 20, 2627. [Google Scholar] [CrossRef] [Green Version]
- Boycheva, S.; Daviet, L.; Wolfender, J.L.; Fitzpatrick, T.B. The rise of operon-like gene clusters in plants. Trends Plant Sci. 2014, 19, 447–459. [Google Scholar] [CrossRef]
- Olsen, K.M.; Small, L.L. Micro-and macroevolutionary adaptation through repeated loss of a complete metabolic pathway. New Phytol. 2018, 219, 757–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Qiu, J.; Li, L.-F.; Lu, B.; Olsen, K.; Fan, L. Genomic clues for crop-weed interactions and evolution. Trends Plant Sci. 2018, 23, 1102–1115. [Google Scholar] [CrossRef] [PubMed]
- Frey, M.; Chomet, P.; Glawischnig, E.; Stettner, C.; Grun, S.; Winklmair, A.; Eisenreich, W.; Bacher, A.; Meeley, R.B.; Briggs, S.P.; et al. Analysis of a chemical plant defense mechanism in grasses. Science 1997, 277, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Bakht, S.; Leggett, M.; Maxwell, C.; Melton, R.; Osbourn, A. A gene cluster for secondary metabolism in oat: Implications for the evolution of metabolic diversity in plants. Proc. Natl. Acad. Sci. USA 2004, 101, 8233–8238. [Google Scholar] [CrossRef] [Green Version]
- Field, B.; Osbourn, A.E. Metabolic diversification—Independent assembly of operon-like gene clusters in different plants. Science 2008, 320, 543–547. [Google Scholar] [CrossRef]
- Field, B.; Fiston-Lavier, A.S.; Kemen, A.; Geisler, K.; Quesneville, H.; Osbourn, A.E. Formation of plant metabolic gene clusters within dynamic chromosomal regions. Proc. Natl. Acad. Sci. USA 2011, 108, 16116–16121. [Google Scholar] [CrossRef]
- Shang, Y.; Ma, Y.; Zhou, Y.; Zhang, H.; Duan, L.; Chen, H.; Zeng, J.; Zhou, Q.; Wang, S.; Gu, W.; et al. Biosynthesis, regulation, and domestication of bitterness in cucumber. Science 2014, 346, 1084–1088. [Google Scholar] [CrossRef]
- Swaminathan, S.; Morrone, D.; Wang, Q.; Fulton, D.B.; Peters, R.J. CYP76M7 is an ent-cassadiene C11α-hydroxylase defining a second multifunctional diterpenoid biosynthetic gene cluster in rice. Plant Cell 2009, 21, 3315–3325. [Google Scholar] [CrossRef] [Green Version]
- Shimura, K.; Okada, A.; Okada, K.; Jikumaru, Y.; Ko, K.W.; Toyomasu, T.; Sassa, T.; Hasegawa, M.; Kodama, O.; Shibuya, N.; et al. Identification of a biosynthetic gene cluster in rice for momilactones. J. Biol. Chem. 2007, 282, 34013–34018. [Google Scholar] [CrossRef] [Green Version]
- Toyomasu, T.; Shenton, M.R.; Okada, K. Evolution of labdane-related diterpene synthases in cereals. Plant Cell Physiol. 2020, 61, 1850–1859. [Google Scholar] [CrossRef]
- Wu, Y.; Hillwig, M.L.; Wang, Q.; Peters, R.J. Parsing a multifunctional biosynthetic gene cluster from rice: Biochemical characterization of CYP71Z6 & 7. FEBS Lett. 2011, 585, 3446–3451. [Google Scholar] [PubMed] [Green Version]
- Wu, Y.; Wang, Q.; Hillwig, M.L.; Peters, R.J. Picking sides: Distinct roles for CYP76M6 and -8 in rice oryzalexin biosynthesis. Biochem. J. 2013, 454, 209–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Hillwig, M.L.; Okada, K.; Yamazaki, K.; Wu, Y.; Swaminathan, S.; Yamane, H.; Peters, R.J. Characterization of CYP76M5–8 indicates metabolic plasticity within a plant biosynthetic gene cluster. J. Biol. Chem. 2012, 287, 6159–6168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Hu, Y.; Akashi, S.; Nojiri, H.; Guo, L.; Ye, C.Y.; Zhu, Q.-H.; Okada, K.; Fan, L. Lateral transfers lead to the birth of momilactone biosynthetic gene clusters in grass. Plant J. 2022, 111, 1354–1367. [Google Scholar] [CrossRef]
- Li, R.; Zhang, J.; Li, Z.; Peters, R.J.; Yang, B. Dissecting the labdane-related diterpenoid biosynthetic gene clusters in rice reveals directional cross-cluster phytotoxicity. New Phytol. 2022, 233, 878–889. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Peters, R.J. The role of momilactones in rice allelopathy. J. Chem. Ecol. 2013, 39, 175–185. [Google Scholar] [CrossRef]
- De La Peña, R.; Sattely, E.S. Rerouting plant terpene biosynthesis enables momilactone pathway elucidation. Nat. Chem. Biol. 2021, 17, 205–212. [Google Scholar] [CrossRef]
- Xu, M.; Hillwig, M.L.; Prisic, S.; Coates, R.M.; Peters, R.J. Functional identification of rice syn-copalyl diphosphate synthase and its role in initiating biosynthesis of diterpenoid phytoalexin/allelopathic natural products. Plant J. 2004, 39, 309–318. [Google Scholar] [CrossRef]
- Otomo, K.; Kanno, Y.; Motegi, A.; Kenmoku, H.; Yamane, H.; Mitsuhashi, W.; Oikawa, H.; Tohima, H.; Itoh, H.; Matsuoka, M.; et al. Diterpene cyclases responsible for the biosynthesis of phytoalexins, momilactones A, B, and oryzalexins A–F in rice. Biosci. Biotechnol. Biochem. 2004, 68, 2001–2006. [Google Scholar] [CrossRef]
- Otomo, K.; Kenmoku, H.; Oikawa, H.; König, W.A.; Toshima, H.; Mitsuhashi, W.; Yamane, H.; Sassa, T.; Toyomasu, T. Biological functions of ent- and syn-copalyl diphosphate synthases in rice: Key enzymes for the branch point of gibberellin and phytoalexin biosynthesis. Plant J. 2004, 39, 886–893. [Google Scholar] [CrossRef]
- Wilderman, P.R.; Xu, M.; Jin, Y.; Coates, R.M.; Peters, R.J. Identification of syn-pimara-7,15-diene synthase reveals functional clustering of terpene synthases involved in rice phytoalexin/allelochemical biosynthesis. Plant Physiol. 2004, 135, 2098–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, L.; Kawaide, H.; Higuchi, T.; Chen, M.; Miyamoto, K.; Hirata, Y.; Kimura, H.; Miyazaki, S.; Teruya, M.; Fujiwara, K.; et al. Genomic evidence for convergent evolution of gene clusters for momilactone biosynthesis in land plants. Proc. Natl. Acad. Sci. USA 2020, 117, 12472–12480. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Hillwig, M.L.; Peters, R.J. CYP99A3: Functional identification of a diterpene oxidase from the momilactone biosynthetic gene cluster in rice. Plant J. 2011, 65, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Kitaoka, N.; Wu, Y.; Zi, J.; Peters, R.J. Investigating inducible short-chain alcohol dehydrogenases/reductases clarifies rice oryzalexin biosynthesis. Plant J. 2016, 88, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Cho, E.M.; Okada, A.; Kenmoku, H.; Otomo, K.; Toyomasu, T.; Mitsuhashi, W.; Sassa, T.; Yajima, A.; Yabuta, G.; Mori, K.; et al. Molecular cloning and characterization of a cDNA encoding ent-cassa-12,15-diene synthase, a putative diterpenoid phytoalexin biosynthetic enzyme, from suspension-cultured rice cells treated with a chitin elicitor. Plant J. 2004, 37, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Prisic, S.; Xu, M.; Wilderman, P.R.; Peters, R.J. Rice contains two disparate ent-copalyl diphosphate synthases with distinct metabolic functions. Plant Physiol. 2004, 136, 4228–4236. [Google Scholar] [CrossRef]
- Ye, Z.; Yamazaki, K.; Minoda, H.; Miyamoto, K.; Miyazaki, S.; Kawaide, H.; Yajima, A.; Nojiri, H.; Yamane, H.; Okada, K. In planta functions of cytochrome P450 monooxygenase genes in the phytocassane biosynthetic gene cluster on rice chromosome 2. Biosci. Biotechnol. Biochem. 2018, 82, 1021–1030. [Google Scholar] [CrossRef] [Green Version]
- Yan, N.; Du, Y.; Liu, X.; Chu, M.; Shi, J.; Zhang, H.; Liu, Y.; Zhang, Z. A comparative UHPLC-QqQ-MS-based metabolomics approach for evaluating Chinese and North American wild rice. Food Chem. 2019, 275, 618–627. [Google Scholar] [CrossRef]
- Shimizu, T.; Lin, F.; Hasegawa, M.; Okada, K.; Nojiri, H.; Yamane, H. Purification and identification of naringenin 7-O methyltransferase, a key enzyme in biosynthesis of flavonoid phytoalexin sakuranetin in rice. J. Biol. Chem. 2012, 287, 19315–19325. [Google Scholar] [CrossRef] [Green Version]
- Bassard, J.E.; Ullmann, P.; Bernier, F.; Werck-Reichhart, D. Phenolamides: Bridging polyamines to the phenolic metabolism. Phytochemistry 2010, 71, 1808–1824. [Google Scholar] [CrossRef]
- Schenck, C.A.; Maeda, H.A. Tyrosine biosynthesis, metabolism, and catabolism in plants. Phytochemistry 2018, 149, 82–102. [Google Scholar] [CrossRef] [PubMed]
- Germain, J.; Deslongchamps, P. Total synthesis of (±)-momilactone A. J. Org. Chem. 2002, 67, 5269–5278. [Google Scholar] [CrossRef] [PubMed]
- Yajima, A.; Mori, K. Diterpenoid total synthesis, XXXII. Synthesis and Absolute Configuration of (−)-Phytocassane D, a Diterpene Phytoalexin Isolated from the Rice Plant, Oryza sativa. Eur. J. Org. Chem. 2000, 24, 4079–4091. [Google Scholar] [CrossRef]
- Mori, K.; Waku, M. Synthesis of oryzalexins A, B and C, the diterpenoidal phytoalexins isolated from rice blast leaves infected with Pyricularia oryzae. Tetrahedron 1985, 41, 5653–5660. [Google Scholar] [CrossRef]
- Jerz, G.; Waibel, R.; Achenbach, H. Cyclohexanoid protoflavanones from the stem-bark and roots of Ongokea gore. Phytochemistry 2005, 66, 1698–1706. [Google Scholar] [CrossRef]
- Ferreira, R.J.; Gajd’acs, M.; Kincses, A.; Spengler, G.; dos Santos, D.J.V.A.; Ferreira, M.-J.U. Nitrogen-containing naringenin derivatives for reversing multidrug resistance in cancer. Bioorg. Med. Chem. 2020, 28, 115798. [Google Scholar] [CrossRef]
- Mizuno, M.; Kojima, H.; Tanaka, T.; Iinuma, M.; Kimura, R.; Zhi-Da, M.; Murata, H. Phenolic constituents from seeds of Coptis Japonica Var. Dissecta. Phytochemistry 1987, 26, 2071–2074. [Google Scholar]
- Aida, Y.; Tamogami, S.; Kodama, O.; Tsukiboshi, T. Synthesis of 7-methoxyapigeninidin and its fungicidal activity against Gloeocercospora sorghi. Biosci. Biotechnol. Biochem. 1996, 60, 1495–1496. [Google Scholar] [CrossRef]
- Oyama, K.; Kondo, T. Total Synthesis of Flavocommelin, a Component of the Blue Supramolecular Pigment from Commelina communis, on the Basis of Direct 6-C-Glycosylation of Flavan. J. Org. Chem. 2004, 69, 5240–5246. [Google Scholar] [CrossRef]
- Selenski, C.; Pettus, T.R.R. (±)-Diinsininone: Made nature’s way. Tetrahedron 2006, 62, 5298–5307. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Park, K.-S.; Lee, C.; Chong, Y. Synthesis of a complete series of O-methyl analogues of naringenin and apigenin. Bull. Korean Chem. Soc. 2007, 28, 2527–2530. [Google Scholar]
- Yamashita, Y.; Hanaya, K.; Shoji, M.; Sugai, T. Simple synthesis of sakuranetin and selinone via a common intermediate, utilizing complementary regioselectivity in the deacetylation of naringenin triacetate. Chem. Pharm. Bull. 2016, 64, 961–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stec, K.; Kozłowska, J.; Wróblewska-Kurdyk, A.; Kordan, B.; Anioł, M.; Gabryś, B. Effect of Naringenin and Its Derivatives on the Probing Behavior of Myzus persicae (Sulz.). Molecules 2020, 25, 3185. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.-H.; Vo, T.-T.-H.; Vo, T.-Q.-N.; Cao, T.-C.-N.; Tran, T.-S. Synthesis and evaluation of the acetylcholinesterase inhibitory activities of some flavonoids derived from naringenin. Sci. World J. 2021, 2021, 4817900. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, M.; Liu, Q.; Huang, J.; Chen, Y. A synthetic view on momilactones and related 9β-H pimarane skeleton diterpenoids. Front. Chem. 2022, 10, 882404. [Google Scholar] [CrossRef]
- Kato, T.; Tsunakawa, M.; Sasaki, N.; Aizawa, H.; Fujita, K.; Kitahara, Y.; Takahashi, N. Growth and germination inhibitors in rice husks. Phytochemistry 1977, 16, 45–48. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T. Assessment of allelopathic potential of root exudate of rice seedlings. Biol. Plant. 2001, 44, 635–638. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T. Rice seedlings release momilactone B into the environment. Phytochemistry 2003, 63, 551–554. [Google Scholar] [CrossRef]
- Chung, I.M.; Hahn, S.J.; Ahmad, A. Confirmation of potential herbicidal agents in hulls of rice, Oryza sativa. J. Chem. Ecol. 2005, 31, 1339–1352. [Google Scholar] [CrossRef]
- Dilday, R.H.; Lin, J.; Yan, W. Identification of allelopathy in the USDA-ARS rice germplasm collection. Aust. J. Exp. Agric. 1994, 34, 907–910. [Google Scholar] [CrossRef]
- Kong, C.H.; Li, H.B.; Hu, F.; Xu, X.H.; Wang, P. Allelochemicals released by rice roots and residues in soil. Plant Soil. 2006, 288, 47–56. [Google Scholar] [CrossRef]
- Chung, I.M.; Kim, J.T.; Kim, S.H. Evaluation of allelopathic potential and quantification of momilactone A, B from rice hull extracts and assessment of inhibitory bioactivity on paddy field weeds. J. Agric. Food Chem. 2006, 54, 2527–2536. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Hasegawa, M.; Ino, T.; Ota, K.; Kujime, H. Contribution of momilactone A and B to rice allelopathy. J. Plant Physiol. 2010, 167, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H. Barnyard grass-induced rice allelopathy and momilactone B. J. Plant Physiol. 2011, 168, 1016–1020. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. The chemical cross talk between rice and barnyardgrass. Plant Signal. Behav. 2011, 6, 1207–1209. [Google Scholar] [CrossRef] [Green Version]
- Mennan, H.; Ngouajio, M.; Sahin, M.; Isik, D.; Altop, E.K. Quantification of momilactone B in rice hulls and the phytotoxic potential of rice extracts on the seed germination of Alisma plantago-aquatica. Weed Biol. Manag. 2012, 12, 29–39. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ota, K. Biological activities of rice allelochemicals momilactone A and B. J. Rice Res. 2013, 1, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Kato-Noguchi, H.; Ino, T. The chemical-mediated allelopathic interaction between rice and barnyard grass. Plant Soil 2013, 370, 267–275. [Google Scholar] [CrossRef]
- Quan, N.V.; Xuan, T.D.; Tran, H.-D.; Dieu Thuy, N.T. Inhibitory Activities of Momilactones A, B, E, and 7-Ketostigmasterol Isolated from Rice Husk on Paddy and Invasive Weeds. Plants 2019, 8, 159. [Google Scholar] [CrossRef] [Green Version]
- Chung, I.M.; Ali, M.; Hahn, S.J.; Siddiqui, N.A.; Lim, Y.H.; Ahmad, A. Chemical constituents from the hulls of Oryza sativa with cytotoxic activity. Chem. Nat. Compd. 2005, 41, 182–189. [Google Scholar] [CrossRef]
- Kim, S.; Park, H.; Park, E.; Lee, S. Cytotoxic and antitumor activity of momilactone B from rice hulls. J. Agric. Food Chem. 2007, 55, 1702–1706. [Google Scholar] [CrossRef]
- Joung, Y.H.; Lim, E.J.; Kim, M.S.; Lim, S.D.; Yoon, S.Y.; Lim, Y.C.; Yoo, Y.B.; Ye, S.K.; Park, T.; Chung, I.M.; et al. Enhancement of hypoxia-induced apoptosis of human breast cancer cells via STAT5b by momilactone B. Int. J. Oncol. 2008, 33, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.C.; Chung, I.-M.; Jin, Y.J.; Song, Y.S.; Seo, S.Y.; Park, B.S.; Cho, K.H.; Yoo, K.S.; Kim, T.-H.; Yee, S.-B.; et al. Momilactone B, an allelochemical of rice hulls, induces apoptosis on human lymphoma cells (Jurkat) in a micromolar concentration. Nutr. Canc. 2008, 60, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Jeong, N.Y.; Kim, G.-Y.; Han, M.H.; Chung, I.-M.; Kim, W.-J.; Yoo, Y.H.; Choi, Y.H. Momilactone B induces apoptosis and G1 arrest of the cell cycle in human monocytic leukemia U937 cells through downregulation of pRB phosphorylation and induction of the cyclin-dependent kinase inhibitor p21Waf1/Cip1. Oncol. Rep. 2014, 31, 1653–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anh, L.H.; Lam, V.Q.; Takami, A.; Khanh, T.D.; Quan, N.V.; Xuan, T.D. Cytotoxic Mechanism of Momilactones A and B against Acute Promyelocytic Leukemia and Multiple Myeloma Cell Lines. Cancers 2022, 14, 4848. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.D.; Minh, T.N.; Anh, L.H.; Khanh, T.D. Allelopathic momilactones A and B are implied in rice drought and salinity tolerance, not weed resistance. Agron. Sustain. Dev. 2016, 36, 52. [Google Scholar] [CrossRef]
- Quan, N.T.; Xuan, T.D. Foliar application of vanillic and p-hydroxybenzoic acids enhanced drought tolerance and formation of phytoalexin momilactones in rice. Arch. Agron. Soil Sci. 2018, 64, 1831–1846. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrao, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Xuan, T.D.; Huong, C.T.; Quan, N.V.; Anh, L.H.; Khanh, T.D.; Rayee, R. Improvement of Salinity Tolerance in Rice Seedlings by Exogenous Magnesium Sulfate Application. Soil Syst. 2022, 6, 69. [Google Scholar] [CrossRef]
- Hoang Anh, L.; Van Quan, N.; Quang Lam, V.; Takami, A.; Dang Khanh, T.; Dang Xuan, T. Rice Momilactones and Phenolics: Expression of Relevant Biosynthetic Genes in Response to UV and Chilling Stresses. Agronomy 2022, 12, 1731. [Google Scholar] [CrossRef]
- Quan, N.V.; Tran, H.D.; Xuan, T.D.; Ahmad, A.; Dat, T.D.; Khanh, T.D.; Teschke, R. Momilactones A and B are α-amylase and α-glucosidase inhibitors. Molecules 2019, 24, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, N.V.; Xuan, T.D.; Tran, H.D.; Ahmad, A.; Khanh, T.D.; Dat, T.D. Contribution of momilactones A and B to diabetes inhibitory potential of rice bran: Evidence from in vitro assays. Saudi Pharm. J. 2019, 27, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Quan, N.V.; Thien, D.D.; Khanh, T.D.; Tran, H.D.; Xuan, T.D. Momilactones A, B, and tricin in rice grain and by-products are potential skin aging inhibitors. Foods 2019, 8, 602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekido, H.; Akatsuka, T. Mode of Action of Oryzalexin D against Pyricularia oryzae. Agric. Biol. Chem. 1987, 51, 1967–1971. [Google Scholar] [CrossRef]
- Sekido, H.; Kamada, K.; Kodama, O.; Akatsuka, T. Antifungal activity of enantiomers of oryzalexins against Pyricularia oryzae. Agric. Biol. Chem. 1987, 51, 2017–2018. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Soytong, K.; Kanokmedhakul, S.; Kanokmedhakul, K.; Poeaim, S. Antifungal activity of microbial nanoparticles derived from Chaetomium spp. against Magnaporthe oryzae causing rice blast. Plant Prot. Sci. 2020, 56, 180–190. [Google Scholar] [CrossRef]
- Jain, S.; Das, A. Virtual screening of natural compounds as combinatorial agents from Indian medicinal plants against estrogen positive breast cancer. Int. J. Incl. Educ. 2020, 3, 266–275. [Google Scholar]
- Zhang, J.; Li, R.; Xu, M.; Hoffmann, R.I.; Zhang, Y.; Liu, B.; Zhang, M.; Yang, B.; Li, Z.; Peters, R.J. A (conditional) role for labdane-related diterpenoid natural products in rice stomatal closure. New Phytol. 2021, 230, 698–709. [Google Scholar] [CrossRef]
- Xu, M.; Galhano, R.; Wiemann, P.; Bueno, E.; Tiernan, M.; Wu, W.; Chung, I.-M.; Gershenzon, J.; Tudzynski, B.; Sesma, A.; et al. Genetic evidence for natural product-mediated plant–plant allelopathy in rice (Oryza sativa). New Phytol. 2012, 193, 570–575. [Google Scholar] [CrossRef]
- Gajbar, T.D.; Satapute, P.; Jogaiah, S. Influence of gamma irradiation on the isolation of bioactive 4-hydroxyisoluceine compound from fenugreek and its enhanced antifungal properties. Physiol. Mol. Plant Pathol. 2022, 118, 101800. [Google Scholar] [CrossRef]
- Shirke, M.D.; Mahesh, H.B.; Gowda, M. Genome-wide comparison of Magnaporthe species reveals a host-specific pattern of secretory proteins and transposable elements. PLoS ONE 2016, 11, e0162458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Kashyap, P.L.; Mahapatra, S.; Jasrotia, P.; Singh, G.P. New and emerging technologies for detecting Magnaporthe oryzae causing blast disease in crop plants. Crop. Prot. 2021, 143, 105473. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T. Concentration and release level of momilactone B in the seedlings of eight rice cultivars. J. Plant Physiol. 2005, 162, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Ino, T. Possible involvement of momilactone B in rice allelopathy. J. Plant Physiol. 2005, 162, 718–721. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H.; Ota, K.; Kujime, H.; Ogawa, M. Effects of momilactone on the protein expression in Arabidopsis germination. Weed Biol. Manag. 2013, 13, 19–23. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ota, K.; Kujime, H. Absorption of momilactone A and B by Arabidopsis thaliana L. and the growth inhibitory effects. J. Plant Physiol. 2012, 169, 1471–1476. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kitajima, S. Momilactone sensitive proteins in Arabidopsis thaliana. Nat. Prod. Commun. 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Sharma, I.; Thakur, A.; Sharma, A.; Singh, N.; Kumar, R.; Sharma, A. Phytoalexins: Implications in Plant Defense and Human Health. In Plant Secondary Metabolites; Springer: Singapore, 2022; pp. 329–353. [Google Scholar]
- Bae, S.; Han, J.W.; Dang, Q.L.; Kim, H.; Choi, G.J. Plant Disease Control Efficacy of Platycladus orientalis and Its Antifungal Compounds. Plants 2021, 10, 1496. [Google Scholar] [CrossRef]
- Reveglia, P.; Cimmino, A.; Masi, M.; Nocera, P.; Berova, N.; Ellestad, G.; Evidente, A. Pimarane diterpenes: Natural source, stereochemical configuration, and biological activity. Chirality 2018, 30, 1115–1134. [Google Scholar] [CrossRef]
- Schmelz, E.A.; Huffaker, A.; Sims, J.W.; Christensen, S.A.; Lu, X.; Okada, K.; Peters, R.J. Biosynthesis, elicitation and roles of monocot terpenoid phytoalexins. Plant J. 2014, 79, 659–678. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.; Wang, Z.; Yang, X.; Guo, L.; Qiu, D.; Zeng, H. Transcriptional profiling of rice treated with MoHrip1 reveal the function of protein elicitor in enhancement of disease resistance and plant growth. Front. Plant Sci. 2016, 7, 1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, A.; Yoshioka, A.; Kariya, K.; Ube, N.; Ueno, K.; Tebayashi, S.; Osaki-Oka, K.; Ishihara, A. Sugars in an aqueous extract of the spent substrate of the mushroom Hypsizygus marmoreus induce defense responses in rice. Biosci. Biotechnol. Biochem. 2021, 85, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, A.; Ando, K.; Yoshioka, A.; Murata, K.; Kokubo, Y.; Morimoto, N.; Ube, N.; Yabuta, Y.; Ueno, M.; Tebayashi, S. Induction of defense responses by extracts of spent mushroom substrates in rice. J. Pestic. Sci. 2019, 44, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Tang, W.; Jiang, L.; Hou, Y.; Yang, F.; Chen, W.; Li, X. Elicitor activity of algino-oligosaccharide and its potential application in protection of rice plant (Oryza saliva L.) against Magnaporthe grisea. Biotechnol. Biotechnol. Equip. 2015, 29, 646–652. [Google Scholar] [CrossRef] [Green Version]
- Yamada, A.; Shibuya, N.; Kodama, O.; Akatsuka, T. Induction of phytoalexin formation in suspension-cultured rice cells by N-Acetylchitooligosaccharides. Biosci. Biotechnol. Biochem. 1993, 57, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, Y.; Oono, Y.; Kanamori, H.; Matsumoto, T.; Itoh, T.; Minami, E. Simultaneous RNA-seq analysis of a mixed transcriptome of rice and blast fungus interaction. PLoS ONE 2012, 7, e49423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dash, M.; Somvanshi, V.S.; Budhwar, R.; Godwin, J.; Shukla, R.N.; Rao, U. A Rice Root-knot nematode Meloidogyne graminicola-resistant mutant rice line shows early expression of plant-defence genes. Planta 2021, 253, 108. [Google Scholar] [CrossRef]
- Daw, B.D.; Zhang, L.H.; Wang, Z.Z. Salicylic acid enhances antifungal resistance to Magnaporthe grisea in rice plants. Australas. Plant Pathol. 2008, 37, 637–644. [Google Scholar]
- Kodama, O.; Suzuki, T.; Miyakawa, J.; Akatsuka, T. Ultraviolet-induced accumulation of phytoalexins in rice leaves. Agric. Biol. Chem. 1988, 52, 2469–2473. [Google Scholar]
- Hasegawa, M.; Mitsuhara, I.; Seo, S.; Imai, T.; Koga, J.; Okada, K.; Yamane, H.; Ohashi, Y. Phytoalexin accumulation in the interaction between rice and the blast fungus. Mol. Plant Microbe Interact. 2010, 23, 1000–1011. [Google Scholar] [CrossRef] [Green Version]
- Kurusu, T.; Hamada, J.; Nokajima, H.; Kitagawa, Y.; Kiyoduka, M.; Takahashi, A.; Hanamata, S.; Ohno, R.; Hayashi, T.; Okada, K.; et al. Regulation of microbe-associated molecular pattern-induced hypersensitive cell death, phytoalexin production, and defense gene expression by calcineurin B-like protein-interacting protein kinases, OsCIPK14/15, in rice cultured cells. Plant Physiol. 2010, 153, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Hamada, H.; Kurusu, T.; Okuma, E.; Nokajima, H.; Kiyoduka, M.; Koyano, T.; Sugiyama, Y.; Okada, K.; Koga, J.; Saji, H.; et al. Regulation of a proteinaceous elicitor-induced Ca2+ influx and production of phytoalexins by a putative voltage-gated cation channel, OsTPC1, in cultured rice cells. J. Biol. Chem. 2012, 287, 9931–9939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umemura, K.; Ogawa, N.; Yamauchi, T.; Iwata, M.; Shimura, M.; Koga, J. Cerebroside elicitors found in diverse phytopathogens activate defense responses in rice plants. Plant Cell Physiol. 2000, 41, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Koga, J.; Yamauchi, T.; Shimura, M.; Ogawa, N.; Oshima, K.; Umemura, K.; Kikuchi, M.; Ogasawara, N. Cerebrosides A and C, sphingolipid elicitors of hypersensitive cell death and phytoalexin accumulation in rice plants. J. Biol. Chem. 1998, 273, 31985–31991. [Google Scholar] [CrossRef] [Green Version]
- Koga, J.; Kubota, H.; Gomi, S.; Umemura, K.; Ohnishi, M.; Kono, T. Cholic acid, a bile acid elicitor of hypersensitive cell death, pathogenesis-related protein synthesis, and phytoalexin accumulation in rice. Plant Physiol. 2006, 140, 1475–1483. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Jikumaru, Y.; Okada, A.; Okada, K.; Koga, J.; Umemura, K.; Minami, E.; Shibuya, N.; Hasegawa, M.; Kodama, O.; et al. Effects of a bile acid elicitor, cholic acid, on the biosynthesis of diterpenoid phytoalexins in suspension-cultured rice cells. Phytochemistry 2008, 69, 973–981. [Google Scholar] [CrossRef]
- Zang, H.; Xie, S.; Zhu, B.; Yang, X.; Gu, C.; Hu, B.; Gao, T.; Chen, Y.; Gao, X. Mannan oligosaccharides trigger multiple defence responses in rice and tobacco as a novel danger-associated molecular pattern. Molecular Plant Pathol. 2019, 20, 1067–1079. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, M.; Kinoshita, H.; Okuno, Y. Antimutagenic activity of sakuranetin from Prunus jamasakura. J. Food Sci. 2003, 68, 52–56. [Google Scholar] [CrossRef]
- Ugocsai, K.; Varga, A.; Molnar, P.; Antus, S.; Molnar, J. Effects of selected flavonoids and carotenoids on drug accumulation and apoptosis induction in multidrug-resistant colon cancer cells expressing MDR1/LRP. In Vivo 2005, 19, 433–438. [Google Scholar]
- Drira, R.; Sakamoto, K. Sakuranetin induces melanogenesis in B16BL6 melanoma cells through inhibition of ERK and PI3K/AKT signaling pathways. Phytother. Res. 2016, 30, 997–1002. [Google Scholar] [CrossRef]
- Hernández, V.; Recio, M.C.; Máñez, S.; Giner, R.M.; Ríos, J.L. Effects of naturally occurring dihydroflavonols from Inula viscosa on inflammation and enzymes involved in the arachidonic acid metabolism. Life Sci. 2007, 81, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Abe, D.; Sekiya, K. Sakuranetin induces adipopenesis of 3T3-L1 cells through enhanced expression of PPARγ2. Biochem. Biophys. Res. Commun. 2008, 372, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Sakoda, C.P.P.; de Toledo, A.C.; Perini, A.; Pinheiro, N.M.; Hiyane, M.I.; dos Grecco, S.S.; de Fátima Lopes Calvo Tibério, I.; Câmara, N.O.S.; de Arruda Martins, M.; Lago, J.H.G.; et al. Sakuranetin reverses vascular peribronchial and lung parenchyma remodeling in a murine model of chronic allergic pulmonary inflammation. Acta Histochem. 2016, 118, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kong, Y.; Wu, D.; Zhang, H.; Wu, J.; Chen, J.; Ding, J.; Hu, L.; Jiang, H.; Shen, X. Three flavonoids targeting the β-hydroxyacyl-acyl carrier protein dehydratase from Helicobacter pylori: Crystal structure characterization with enzymatic inhibition assay. Protein Sci. 2008, 17, 1971–1978. [Google Scholar] [CrossRef] [Green Version]
- Grecco, S.S.; Reimao, J.Q.; Tempone, A.G.; Sartorelli, P.; Cunha, R.L.; Romoff, P.; Ferreira, M.J.P.; Favero, O.A.; Lago, J.H.G. In vitro antileishmanial and antitrypanosomal activities of flavanones from Baccharis retusa DC (Asteraceae). Exp. Parasitol. 2012, 130, 141–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moulishankar, A.; Lakshmanan, K. Data on molecular docking of naturally occurring flavonoids with biologically important targets. Data Brief 2020, 29, 105243. [Google Scholar] [CrossRef]
- Máñez, S.; Hernández, V.; Giner, R.M.; Ríos, J.L.; del Carmen Recio, M. Inhibition of pro-inflammatory enzymes by inuviscolide, a sesquiterpene lactone from Inula viscosa. Fitoterapia 2007, 78, 329–331. [Google Scholar] [CrossRef]
- Hernández, V.; del Carmen Recio, M.; Máñez, S.; Prieto, J.M.; Giner, R.M.; Ríos, J.L. A mechanistic approach to the in vivo anti-inflammatory activity of sesquiterpenoid compounds isolated from Inula viscosa. Planta Med. 2001, 67, 726–731. [Google Scholar] [CrossRef]















| Entry | Compound | Skeletal Type | References |
|---|---|---|---|
| 1 | (±)-Momilactone A | (9β-H)-Pimarane | [143] |
| 2 | (−)-Phytocassane D | ent-Cassane | [144] |
| 3 | (+)-Oryzalexin A, B and C | ent-Isopimarane | [145] |
| 4 | (−)-Sakuranetin | Flavanone | [146,147] *, |
| 5 | (±)-Sakuranetin | Flavanone | [148], [149,150] *, [151], [152,153,154,155] * |
| Metabolite/s | Activity | Experimental Model | References |
|---|---|---|---|
| Momilactones | Antimicrobial and allelopathic | Structural characterization of momilactones A and B isolated from rice husk (cv. Koshihikari), which inhibited rice root growth at less than 100 ppm. | [27] |
| Evaluation of inhibitory activity of momilactones A and B isolated from rice (cvs. Koshihikari and Surjamukhi) seed hulls on rice and lettuce seed germination. | [55] | ||
| Structural characterization of momilactone C isolated from rice husk (cv. Koshihikari) and evaluation of inhibitory activity toward germination of lettuce seeds. | [42] | ||
| Increased responsiveness of rice to the infection by M. grisea after treatment of leaves with the resistance inducer WL28325 (2,2-dichloro-3,3-dimethylcyclopropane carboxylic acid) in terms of momilactones A and B synthesis. | [56] | ||
| Evaluation of inhibitory activities of momilactones A and B isolated from rice (cv. Koshihikari) on lettuce seed germination and rice root growth. Bioassays with several semisynthetic momilactone derivatives to establish the functional groups responsible for the bioactivity. | [157] | ||
| Structural characterization of momilactones A and B from coleoptiles of etiolated plantlets of rice (cv. Sasashigure) exposed to UV radiation. Evaluation of anti-fungal activity on Cladosporium cucumerinum through TLC bioassay. | [28] | ||
| Evaluation of inhibitory effect of rice root exudates (cvs. Hinohikari, Nipponbare, Norin 8, Kamenoo, Kinuhikari, Koshihikari, Sasanishiki, and Yukihikari) on co-cultivated seedlings of alfalfa (Medicago sativa L.), cress (Lepidium sativum L.) or lettuce (Lactuca sativa L.). | [158] | ||
| Evaluation of inhibitory activity of momilactone B in rice (cv. Koshihikari) root exudates on growth of cress and lettuce. | [58,159] | ||
| Evaluation of phytotoxicity of momilactones A and B and other putative allelochemicals from rice (cv. Hochokjindo) hulls against duckweed (Lemna paucicostata). Evaluation of inhibitory effects of identified compounds on germination and growth of the three weed species Amaranthus retroflexus, Cyperus difformis, and Leptochloa chinensis. | [160] | ||
| Release of momilactone B and other allelochemicals into the soil promoted by the presence of barnyard grass by two allelopathic rice varieties PI312777 [from USDA-ARS rice germplasm collection [161] and Huagan-1, and the non-allelopathic rice variety Huajingxian. | [162] | ||
| Evaluation of allelopathic potential of rice hull extracts from ninety-nine rice varieties containing momilactones A and B on barnyard grass germination and growth. | [163] | ||
| Evaluation of inhibitory activity of acetone extracts from the moss Calohypnum plumiforme (syn. Hypnum plumaeforme) on the growth of angiosperms (Arabidopsis thaliana, Nicotiana tabacum), mosses (Physcomitrella patens and H. plumaeforme), and a liverwort (Jungermannia subulata cultured cells). | [48] | ||
| Evaluation of herbicidal activity (inhibition of germination and elongation of shoot and root) of momilactones A and B from rice hulls on barnyard grass and monochoria (Monochoria vaginalis). Evaluation of antifungal activity by agar dilution method against Botrytis cinerea, Fusarium solani, F. oxysporum, and Colletrotrichum gloeosporioides. Evaluation of antibacterial activity by disc diffusion method against Pseudomonus ovalis, Bacillus cereus, B. pumilus, and Escherichia coli. | [19] | ||
| Evaluation of growth inhibitory activity (lengths of hypocotyls) of momilactones A and B released in the medium by in vitro-grown rice (cv. Nipponbare) seedlings on lettuce and Chinese cabbage (Brassica rapa). | [50] | ||
| Evaluation of inhibitory activity of momilactone A from root exudates of rice (cv. Koshihikari) on growth of cress (length of root and hypocotyl). | [60,61] | ||
| Elicitation of momilactone accumulation and secretion in C. plumiforme through UV, jasmonic acid, metals (CuCl2 and FeCl2), and a protein phosphatase inhibitor (cantharidin). | [63,64] | ||
| Evaluation of inhibitory activity of H2O:MeOH extracts of soil under colonies of C. plumiforme on growth of cress (Lepidium sativum), lettuce (Lactuca sativa), lucerne (Medicago sativa), ryegrass (Lolium multiflorum), timothy (Phleum pratense), Digitaria sanguinalis, and Echinochloa crus-galli. | [65] | ||
| Evaluation of inhibitory activity of momilactones A and B from root exudates of rice (cv. Koshihikari) on growth of barnyard grass (length of shoot and hypocotyl). Evaluation of allelopathic activity of six rice cvs. (Hinohikari, Kamenoo, Kinuhikari, Koshihikari, Nipponbare, Norin 8, Sasanishiki, and Yukihikari) by donor–receiver bioassay. | [164] | ||
| Evaluation of momilactone B production and release in rice (O. sativa) and barnyard grass (E. crus-galli) co-cultures. | [165,166] | ||
| Screening of 41 different rice cvs. for their ability to reduce germination, root growth, and root dry weight of Alisma plantago-aquatica. Quantification of momilactone B in the studied rice cvs. | [167] | ||
| Evaluation of allelopathic activities of momilactones A and B from husks of rice (cv. Koshihikari) against different plant species, including nine weed species [Cress, lettuce, alfalfa (Medicago sativa), ryegrass (Lolium multiflorum), timothy, barnyard grass, Echinochloa colonum, crabgrass (Digitaria sanguinalis), and Arabidopsis thaliana], and four rice cvs. (Koshihikari, Nipponbare, Norin 8, and Sasanishiki). | [168] | ||
| Evaluation of allelopathic activity of barnyard grass in response to momilactone B from rice (cv. Koshihikari) seedlings or root exudates. | [169] | ||
| Evaluation of inhibitory activity of momilactones A, B, E from rice (cv. Koshihikari) husk on germination rate and root and shoot elongation of lettuce, barnyard grass, and tall goldenrod (Solidago altissima). | [170] | ||
| Anti-cancer | Evaluation of cytotoxic activity of momilactones A and B from rice (cv. Hochokjindo) hulls against P388 murine leukemia cells. | [171] | |
| Evaluation of cytotoxic and antitumor activity of methanolic extract and momilactone B from rice hulls by MTT-dye reduction assay against human colon cancer cells and colonic aberrant crypt foci (ACF) assay in 1,2-dimethylhydrazine (DMH)-injected F344 male rats, respectively. | [172] | ||
| Investigation of the molecular mechanism responsible for the inhibitory effects of momilactone B on the growth of cultured human breast cancer T47D cells. | [173] | ||
| Evaluation of antitumor efficacy by inducing apoptosis in several mammalian blood cancer cells, including human leukemic T cells. | [174] | ||
| Investigation of the molecular mechanism responsible for the inhibitory effects of momilactone B on cultured human leukemia U937 cell growth. | [175] | ||
| Investigation of the molecular mechanism responsible for the cytotoxic activity of momilactones A and B against multiple myeloma U266 and acute promyelocytic leukemia HL-60 cell lines. | [176] | ||
| Increased tolerance to salinity and drought stresses | Correlation between the content of momilactones A and B and tolerance to salinity and drought in 30 rice cultivars. | [177] | |
| Induction of momilactones A and B by application of exogenous vanillic acid and p-hydroxybenzoic acid to two rice cvs., one drought-tolerant (Nep nanh ngua Hai phong), and one drought-susceptible rice (Re nuoc). | [178] | ||
| Enhancement of momilactone B and phenolic acids production by exogenous application of MgSO4 to salinity tolerant (BC15) and salinity susceptible (DT84DB) rice varieties [179]. | [180] | ||
| Increased tolerance to UV and chilling stresses | Induction of expression of genes related to the biosynthesis of momilactones and phenolics and enhancement of their accumulation by UV. | [181] | |
| Anti-diabetic and anti-obesity | Evaluation of in vitro inhibitory activity of momilactones A and B from rice (cv. Koshihikari) hulls on α-amylase and α-glucosidase (in comparison with the known commercial diabetes inhibitors, acarbose, and quercetin). | [182] | |
| Evaluation of in vitro inhibitory activity of momilactones A and B from rice bran on pancreatic α-amylase and α-glucosidase (in comparison with the known diabetes inhibitor γ-oryzanol). | [183] | ||
| Antioxidant | Evaluation of antioxidant activity of momilactones A and B from rice hulls through DPPH radical scavenging capacity assay. | [19] | |
| Evaluation of antioxidant activity of momilactones A and B from rice grains of different cultivars (Koshihikari, Shinnosuke, Seiten no hekireki, Ginga no shizuku, Ho no mai) through ABTS radical cation decolorization assay. | [184] | ||
| Anti-inflammatory | Structural characterization of momilactones D and E from rice (cv. Chucheongbyeo) root and evaluation of their ability to inhibit the production of NO and iNOS mRNA and protein expression in LPS-stimulated RAW264.7 macrophages. | [43] | |
| Anti-aging | Evaluation of the ability of momilactones A and B from rice grains of different cultivars (Koshihikari, Shinnosuke, Seiten no hekireki, Ginga no shizuku, Ho no mai) to relieve wrinkles, skin, and freckles by in vitro enzymatic assays on pancreatic elastase and tyrosinase. | [184] | |
| Oryzalexins A-F | Antimicrobic | Evaluation of inhibitory activity of oryzalexins A, B, and C against M. grisea. | [33] |
| Evaluation of inhibitory activity of oryzalexin D against M. grisea. | [185] | ||
| Evaluation of inhibitory activity of oryzalexin A, B, and C synthetic enantiomers against M. grisea. | [186] | ||
| Evaluation of inhibitory activity of oryzalexins A-F against Magnaporthe grisea. | [20] | ||
| Evaluation of inhibitory activity of oryzalexin B and sakuranetin induced in rice leaves by elicitation with fungal metabolites and nanoparticles on spore production in M. grisea. | [187] | ||
| Anti-cancer | Evaluation of binding potential of a combination of oryzalexin B and other compounds on six potential receptors in estrogen receptor-positive breast cancer. | [188] | |
| Regulation of stomatal closure | Evaluation of stomatal closure and susceptibility to drought in cps2 and cps4 knockout lines. | [189] | |
| Oryzalexin S | Allelopathic | Evaluation of allelopathic activity of oryzalexin S against L. sativa cv. Black-seeded Simpson and E. crus-galli cv. Kudiraivali in O. sativa Oskls4 knockouts. | [190] |
| Antimicrobic | Evaluation of inhibitory activity of oryzalexins S against M. grisea. | [20] | |
| Regulation of stomatal closure | Evaluation of stomatal closure and susceptibility to drought in cps2 and cps4 knockout lines. | [189] | |
| Phytocassanes | Antimicrobial | Evaluation of inhibition of M. grisea spore germination by phytocassanes A-D from rice leaves infected with M. grisea and from rice stems infected with Rhizoctonia solani. | [29] |
| Evaluation of inhibition of M. grisea spore germination by phytocassanes E from rice suspension-cultured cells treated with a mycelial extract of the pathogenic potato fungus Phytophthora infestans. | [30] | ||
| Evaluation of inhibition of M. grisea spore production by phytocassanes A-F from rice leaves elicited by UV light. | [31] |
| Biological Activity | Experimental Model | References |
|---|---|---|
| Antifungal | Magnaporthe grisea (syn. M. oryzae, Pyricularia grisea/oryzae)—Inhibition of spore germination and germ tube growth. | [70] |
| M. grisea—Fungal colony growth inhibition assay. | [95] | |
| Antimutagenic | Salmonella typhimurium TA1535/pSK1002—Suppressive effect on umu gene expression of SOS response against the mutagen AF-2. | [220] |
| Anticancer | Multidrug-resistant Colo 320 human colon cancer cells—Growth inhibition by induction of apoptosis. | [221] |
| Colon carcinoma HCT-116 human cells—Cytotoxicity test through MTT assay. | [18] | |
| B16BL6 mouse melanoma cells—Dose-dependent stimulation of melanogenesis. | [222] | |
| Antiallergic | Blood flow decrease in the tail vein microcirculation of mice subjected to HEL-sensitization as a monitor—Measurement of platelet aggregation of whole blood induced by ADP through WBA analyzer. | [85] |
| Anti-inflammatory | In vivo induction of acute inflammation by topical application of TPA to mouse ears or by subcutaneous injection of PLA2 into mouse paws. In vitro tests based on the effect of sakuranetin and other dihydroflavonols on arachidonic acid metabolism and release and/or activity of enzymes implicated in the inflammatory response like elastase, MPO, and PKC. | [223] |
| Adipogenesis induction—glucose uptake stimulation | Differentiation of 3T3-L1 murine preadipocytes to adipocytes. Expression of genes involved in development of adipocyte phenotypes. Evaluation of basal- and insulin-stimulated glucose uptake. | [224] |
| Anti-inflammatory and antioxidant | Murine model (male BALB/c mice) of chronic allergic pulmonary inflammation—Histopathological analysis: evaluation of extracellular matrix remodeling, inflammation, and oxidative stress in pulmonary vessels and lung parenchyma; quantification of the vascular wall thickness and the VEGF levels. | [225] |
| Anti-Helicobacter pylori | Study of the interaction between sakuranetin and other flavonoids with HpFabZ protein from H. pylori by enzymatic and crystalline structure analyses. | [226] |
| Antileishmanial and antitrypanosomal | In vitro antileishmanial and antitrypanosomal tests. | [227] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valletta, A.; Iozia, L.M.; Fattorini, L.; Leonelli, F. Rice Phytoalexins: Half a Century of Amazing Discoveries; Part I: Distribution, Biosynthesis, Chemical Synthesis, and Biological Activities. Plants 2023, 12, 260. https://doi.org/10.3390/plants12020260
Valletta A, Iozia LM, Fattorini L, Leonelli F. Rice Phytoalexins: Half a Century of Amazing Discoveries; Part I: Distribution, Biosynthesis, Chemical Synthesis, and Biological Activities. Plants. 2023; 12(2):260. https://doi.org/10.3390/plants12020260
Chicago/Turabian StyleValletta, Alessio, Lorenzo Maria Iozia, Laura Fattorini, and Francesca Leonelli. 2023. "Rice Phytoalexins: Half a Century of Amazing Discoveries; Part I: Distribution, Biosynthesis, Chemical Synthesis, and Biological Activities" Plants 12, no. 2: 260. https://doi.org/10.3390/plants12020260