
Protein Profiling of Psittacanthus calyculatus during Mesquite Infection
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. The Study System
2.2. Germination and Infection Development
2.3. Proteomic Dynamics during Invasive Stages
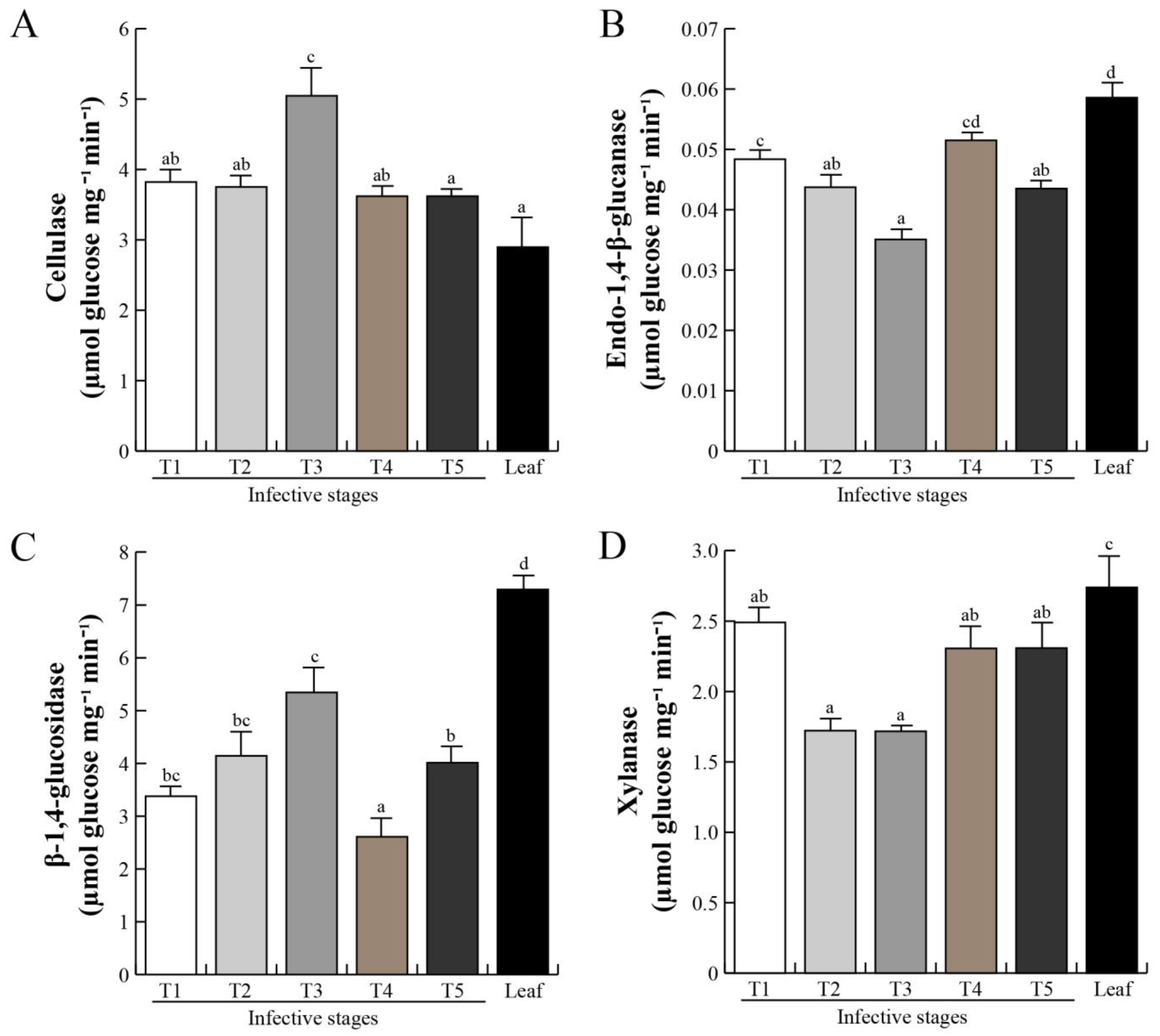
2.4. Cell Wall-Degrading Enzymes during Host Invasion
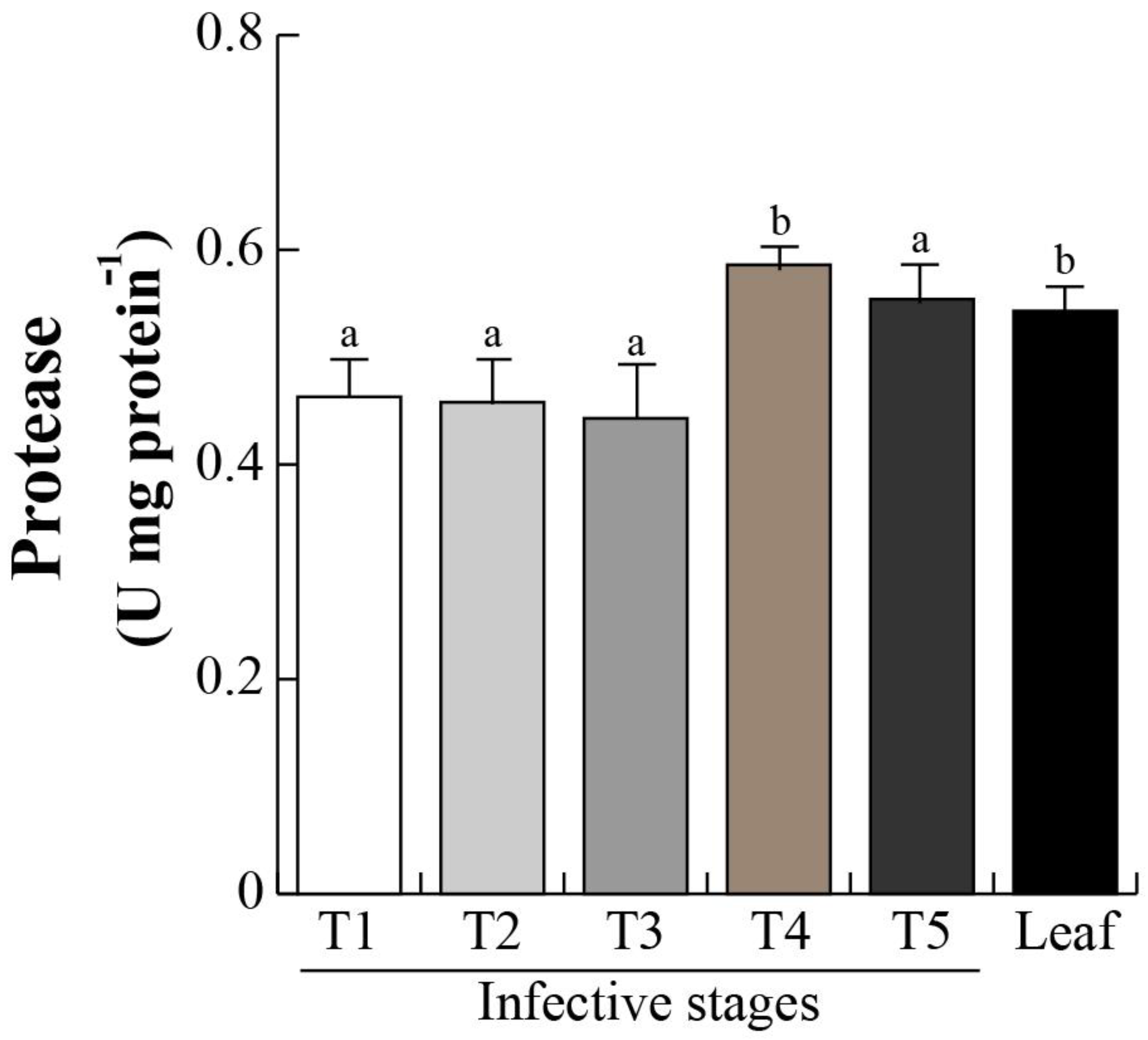
2.5. Protease Activity and Chlorophyll Content
2.6. Phytohormone Level during Invasive Stages
3. Discussion
3.1. Morphological Establishment of Mistletoe Invasive Stages
3.2. Protein Functions Involved in Seed Germination and Haustorium Development
3.3. Spatial Secretion of Cell Wall-Degrading Enzymes
3.4. Metabolic Aspects during Germination
3.5. Phytohormone Regulation of Germination and Seedling Establishment
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Study Site
5.2. Initial Infection Stage Collection
5.3. Total Protein Extraction of P. calyculatus Stages and Leaves
5.4. SDS-PAGE
5.5. Two-Dimensional Electrophoresis (2-DE)
5.6. Protein in Gel Digestion, LC-MS/MS, and Data Analysis
5.7. Cellulase Activity
5.8. Endo-1,4-β-glucanase
5.9. β-glucosidase Activity
5.10. Xylanase Activity
5.11. Proteolytic Activity
5.12. Chlorophyll Content
5.13. Phytohormone Determination
5.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kokla, A.; Melnyk, C.W. Developing a thief: Haustoria formation in parasitic plants. Dev. Biol. 2018, 442, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.M.; McLellan, R.C.; Fontúrbel, F.E. Functional Roles of Parasitic Plants in a Warming World. Annu. Rev. Ecol. Evol. Syst. 2022, 53, 25–45. [Google Scholar] [CrossRef]
- Teixeira-Costa, L.; Davis, C.C. Life history, diversity, and distribution in parasitic flowering plants. Plant Physiol. 2021, 187, 32–51. [Google Scholar] [CrossRef] [PubMed]
- Bian, P.; Sun, C.; Cao, X.; Yao, Z.; Zhang, X.; Zhao, S. Screening of Haustorium Induction Factors of Phelipanche aegyptiaca Pers. Based on Metabolome Analysis of Cucumis melo L. Root Exudates. Agronomy 2023, 13, 128. [Google Scholar] [CrossRef]
- Nickrent, D.L.; Su, H.J.; Lin, R.Z.; Devkota, M.P.; Hu, J.M.; Glatzel, G. Examining the Needle in the Haystack: Evolutionary Relationships in the Mistletoe Genus Loranthus (Loranthaceae). Syst. Bot. 2021, 46, 403–415. [Google Scholar] [CrossRef]
- Nickrent, D.L.; Malécot, V.; Vidal-Russell, R.; Der, J.P. A revised classification of santalales. Taxon 2010, 59, 538–558. [Google Scholar] [CrossRef]
- Nickrent, D.L.; Anderson, F.; Kuijt, J. Inflorescence evolution in Santalales: Integrating morphological characters and molecular phylogenetics. Am. J. Bot. 2019, 106, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Quintana-Rodríguez, E.; Ramírez-Rodríguez, A.G.; Ramírez-Chávez, E.; Molina-Torres, J.; Camacho-Coronel, X.; Esparza-Claudio, J.; Heil, M.; Orona-Tamayo, D. Biochemical traits in the flower lifetime of a mexican mistletoe parasitizing mesquite biomass. Front. Plant Sci. 2018, 9, 1031. [Google Scholar] [CrossRef] [Green Version]
- Hanousek Čiča, K.; Lukin, P.; Derewiaka, D.; Mrvčić, J.; Stanzer, D. Chemical Composition, Physical Properties, and Aroma Profile of Ethanol Macerates of Mistletoe (Viscum album). Beverages 2022, 8, 46. [Google Scholar] [CrossRef]
- Kuijt, J.; Hansen, B. Loranthaceae. In Flowering Plants. Eudicots; Springer: Berlin/Heidelberg, Germany, 2015; pp. 73–119. [Google Scholar]
- Fadini, R.F.; Fischer, E.; Castro, S.J.; Araujo, A.C.; Ornelas, J.F.; de Souza, P.R. Bat and bee pollination in Psittacanthus mistletoes, a genus regarded as exclusively hummingbird-pollinated. Ecology 2018, 99, 1239–1241. [Google Scholar] [CrossRef]
- Mathiasen, R.L.; Nickrent, D.L.; Shaw, D.C.; Watson, D.M. Mistletoes: Pathology, systematics, ecology, and management. Plant Dis. 2008, 92, 988–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agne, M.C.; Shaw, D.C.; Woolley, T.J.; Queijeiro-Bolaños, M.E. Effects of dwarf mistletoe on stand structure of lodgepole pine forests 21-28 years post-mountain pine beetle epidemic in central Oregon. PLoS ONE 2014, 9, e107532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szmidla, H.; Tkaczyk, M.; Plewa, R.; Tarwacki, G.; Sierota, Z. Impact of common mistletoe (Viscum album L.) on Scots pine forests—A call for action. Forests 2019, 10, 847. [Google Scholar] [CrossRef] [Green Version]
- Diniz, U.M.; Fischer, N.L.; Aguiar, L.M. Changing the main course: Strong bat visitation to the ornithophilous mistletoe Psittacanthus robustus (Loranthaceae) in a Neotropical savanna. Biotropica 2022, 54, 478–489. [Google Scholar] [CrossRef]
- Dobrecky, C.B.; Lucangioli, S.E.; Wagner, M.L. The Argentine Mistletoes Ligaria cuneifolia (Ruiz & Pav.) Tiegh (Loranthaceae) and Phoradendron liga (Gillies ex Hook. & Arn.) Eichler (Santalaceae). Thirty Years of Research. Chem. Biodivers. 2022, 19, e202200565. [Google Scholar]
- Kuijt, J. Monograph of Psittacanthus (Loranthaceae). Syst. Bot. Monogr. 2009, 86, 1–361. [Google Scholar]
- Robayo, C.; Marquínez, X.; Raz, L.; Nickrent, D.-L. Floral anatomy of the plant Psittacanthus schiedeanus (Loranthaceae). Rev. De Biol. Trop. 2020, 68, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Azpeitia, F.; Lara, C. Reproductive biology and pollination of the parasitic plant Psittacanthus calyculatus (Loranthaceae) in central México1. J. Torrey Bot. Soc. 2006, 133, 429–438. [Google Scholar] [CrossRef]
- Moustapha, B.; Marina, G.-A.D.; Raúl, F.-O.; Raquel, C.-M.; Mahinda, M. Chemical constituents of the Mexican mistletoe (Psittacanthus calyculatus). Molecules 2011, 16, 9397–9403. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.C.; Guimarães, A.F.; Teodoro, G.S.; Bastos, S.S.; de Castro, E.M.; van den Berg, E. The enemy within: The effects of mistletoe parasitism on infected and uninfected host branches. Plant Ecol. 2021, 222, 639–645. [Google Scholar] [CrossRef]
- Aguilar-Venegas, J.M. La Herramienta de uno Puede ser el Arma del otro: Biomoleculas Involucradas en el Proceso Inicial Infectivo del Muerdago en Arboles de Mezquite. 2019 UNAM. Available online: https://ru.dgb.unam.mx/handle/DGB_UNAM/TES01000794908 (accessed on 12 November 2022).
- Changhu, L. Biology of mistletoe (Viscum coloratum) and its seed dispersal by frugivoro us birds. Acta Ecol. Sin. 2003, 23, 834–839. [Google Scholar]
- McKey, D. The ecology of coevolved seed dispersal systems. In Coevolution of Animals and Plants; University of Texas Press: New York, NY, USA, 1975; pp. 159–191. [Google Scholar]
- Irving, L.J.; Cameron, D.D. You are what you eat: Interactions between root parasitic plants and their hosts. Adv. Bot. Res. 2009, 50, 87–138. [Google Scholar]
- Collazo-Vázquez, I.; Geils, B. Psittacanthus in Mexico; Department of Agriculture, USA, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2002; pp. 9–12. [Google Scholar]
- Teixeira-Costa, L. A living bridge between two enemies: Haustorium structure and evolution across parasitic flowering plants. Braz. J. Bot. 2021, 44, 165–178. [Google Scholar] [CrossRef]
- Tennakoon, K.; Pate, J. Effects of parasitism by a mistletoe on the structure and functioning of branches of its host. Plant Cell Environ. 1996, 19, 517–528. [Google Scholar] [CrossRef]
- Lech, P.; Żółciak, A.; Hildebrand, R. Occurrence of European mistletoe (Viscum album L.) on forest trees in Poland and its dynamics of spread in the period 2008–2018. Forests 2020, 11, 83. [Google Scholar] [CrossRef] [Green Version]
- Albert, M.; Werner, M.; Proksch, P.; Fry, S.; Kaldenhoff, R. The cell wall-modifying xyloglucan endotransglycosylase/hydrolase LeXTH1 is expressed during the defence reaction of tomato against the plant parasite Cuscuta reflexa. Plant Biol. 2004, 6, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.; Krause, K. Activity of xyloglucan endotransglucosylases/hydrolases suggests a role during host invasion by the parasitic plant Cuscuta reflexa. PLoS ONE 2017, 12, e0176754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutuku, J.M.; Cui, S.; Yoshida, S.; Shirasu, K. Orobanchaceae parasite–host interactions. New Phytol. 2021, 230, 46–59. [Google Scholar] [CrossRef]
- Fishman, M.R.; Shirasu, K. How to resist parasitic plants: Pre-and post-attachment strategies. Curr. Opin. Plant Biol. 2021, 62, 102004. [Google Scholar] [CrossRef] [PubMed]
- Clarke, C.R.; Timko, M.P.; Yoder, J.I.; Axtell, M.J.; Westwood, J.H. Molecular dialog between parasitic plants and their hosts. Annu. Rev. Phytopathol. 2019, 57, 279–299. [Google Scholar] [CrossRef]
- Singh, A.; Singh, M. Cell wall degrading enzymes in Orobanche aegyptiaca and its host Brassica campestris. Physiol. Plant. 1993, 89, 177–181. [Google Scholar] [CrossRef]
- Brun, G.; Spallek, T.; Simier, P.; Delavault, P. Molecular actors of seed germination and haustoriogenesis in parasitic weeds. Plant Physiol. 2021, 185, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, B.; Guo, Q.; Song, L.; Chen, L.; Wang, C. Construction of a haustorium development associated SSH library in Thesium chinense and analysis of specific ESTs included by Imperata cylindrica. Biochem. Syst. Ecol. 2016, 64, 46–52. [Google Scholar] [CrossRef]
- Spallek, T.; Melnyk, C.W.; Wakatake, T.; Zhang, J.; Sakamoto, Y.; Kiba, T.; Yoshida, S.; Matsunaga, S.; Sakakibara, H.; Shirasu, K. Interspecies hormonal control of host root morphology by parasitic plants. Proc. Natl. Acad. Sci. USA 2017, 114, 5283–5288. [Google Scholar] [CrossRef]
- Xiao, T.T.; Kirschner, G.K.; Kountche, B.A.; Jamil, M.; Savina, M.; Lube, V.; Mironova, V.; Al Babili, S.; Blilou, I. A PLETHORA/PIN-FORMED/auxin network mediates prehaustorium formation in the parasitic plant Striga hermonthica. Plant Physiol. 2022, 189, 2281–2297. [Google Scholar] [CrossRef]
- Aoki, N.; Cui, S.; Yoshida, S. Cytokinins Induce Prehaustoria Coordinately with Quinone Signals in the Parasitic Plant Striga hermonthica. Plant Cell Physiol. 2022, 63, 1446–1456. [Google Scholar] [CrossRef]
- Muche, M.; Muasya, A.M.; Tsegay, B.A. Biology and resource acquisition of mistletoes, and the defense responses of host plants. Ecol. Process. 2022, 11, 1–13. [Google Scholar] [CrossRef]
- Esquerre-Tugaye, M.T.; Boudart, G.; Dumas, B. Cell wall degrading enzymes, inhibitory proteins, and oligosaccharides participate in the molecular dialogue between plants and pathogens. Plant Physiol. Biochem. 2000, 38, 157–163. [Google Scholar] [CrossRef]
- Jhu, M.-Y.; Sinha, N.R. Parasitic Plants: An Overview of Mechanisms by Which Plants Perceive and Respond to Parasites. Annu. Rev. Plant Biol. 2022, 73, 433–455. [Google Scholar] [CrossRef]
- Ge, D.; Yeo, I.-C.; Shan, L. Knowing me, knowing you: Self and non-self recognition in plant immunity. Essays Biochem. 2022, 66, 447–458. [Google Scholar] [PubMed]
- Ogawa, S.; Wakatake, T.; Spallek, T.; Ishida, J.K.; Sano, R.; Kurata, T.; Demura, T.; Yoshida, S.; Ichihashi, Y.; Schaller, A.; et al. Subtilase activity in intrusive cells mediates haustorium maturation in parasitic plants. Plant Physiol. 2020, 185, 1381–1394. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.C. The mechanism of host-induced germination in root parasitic plants. Plant Physiol. 2021, 185, 1353–1373. [Google Scholar] [CrossRef]
- Geils, B.; Hawksworth, F. Damage, effects, and importance of dwarf mistletoes. In Mistletoes of North American Conifers. Gen. Tech. Rep. RMRS-GTR-98; Geils Brian, W., Cibrián Tovar, J., Moody, B., Eds.; tech. coords; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2002; Volume 98, pp. 57–65. [Google Scholar]
- Ben-Hod, G.; Nun, N.B.; Tzaban, S.; Mayer, A. Inhibitors of polygalacturonase in calli of Orobanche aegyptiaca. Phytochemistry 1997, 45, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.M.; Kwon, Y.K.; Kim, J.H.; Song, I.-J.; Lee, H.-Y.; Choi, D.-W.; Liu, J.R.; Kim, S.W. Transcriptome analysis of mistletoe (Viscum album) haustorium development. Hortic. Environ. Biotechnol. 2014, 55, 352–361. [Google Scholar] [CrossRef]
- Losner-Goshen, D.P.; Portnoy, V.H.; Mayer, A.M.; Joel, D.M. Pectolytic activity by the haustorium of the parasitic PlantOrobancheL.(Orobanchaceae) in host roots. Ann. Bot. 1998, 81, 319–326. [Google Scholar] [CrossRef]
- Ben-Hod, G.; Losner, D.; Joel, D.M.; Mayer, A.M. Pectin methylesterase in calli and germinating seeds of Orobanche aegyptiaca. Phytochemistry 1993, 32, 1399–1402. [Google Scholar] [CrossRef]
- Lin, Y.L.; Chung, C.L.; Huang, P.j.; Chen, C.H.; Fang, S.C. Revised annotation and extended characterizations of components of the Chlamydomonas reinhardtii SUMOylation system. Plant Direct 2020, 4, e00266. [Google Scholar] [CrossRef]
- Negi, S.; Bhakta, S.; Ganapathi, T.; Tak, H. MusaNAC29-like transcription factor improves stress tolerance through modulation of phytohormone content and expression of stress responsive genes. Plant Sci. 2023, 326, 111507. [Google Scholar] [CrossRef]
- Shin, E.J.; Shin, H.M.; Nam, E.; Kim, W.S.; Kim, J.H.; Oh, B.H.; Yun, Y. DeSUMOylating isopeptidase: A second class of SUMO protease. EMBO Rep. 2012, 13, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Ishida, J.K.; Wakatake, T.; Yoshida, S.; Takebayashi, Y.; Kasahara, H.; Wafula, E.; Depamphilis, C.W.; Namba, S.; Shirasu, K. Local auxin biosynthesis mediated by a YUCCA flavin monooxygenase regulates haustorium development in the parasitic plant Phtheirospermum japonicum. Plant Cell 2016, 28, 1795–1814. [Google Scholar] [CrossRef] [Green Version]
- Geils, B.; Collazo, I.V. Loranthaceae and Viscaceae in North America. In Mistletoes of North American Conifers. Gen. Tech. Rep. RMRS-GTR-98; Geils Brian, W., Cibrián Tovar, J., Moody, B., Eds.; tech. coords; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2002; Volume 98, pp. 1–8. [Google Scholar]
- Thoday, D. The haustorial system of Viscum album. J. Exp. Bot. 1951, 2, 1–19. [Google Scholar] [CrossRef]
- Kuijt, J. Parasitism in Pholisma (Lennoaceae). I. External morphology of subterranean organs. Am. J. Bot. 1966, 53, 82–86. [Google Scholar] [CrossRef]
- Heide-Jørgensen, H. Parasitic flowering plants. In Parasitic Flowering Plants; Brill: Leiden, The Netherlands, 2008; Available online: https://brill.com/display/title/15231.
- Rocha, D.; Santhoshkumar, A. Host plant influence on haustorial growth and development of Indian sandalwood (Santalum album). In Indian Sandalwood; Springer: Berlin/Heidelberg, Germany, 2022; pp. 229–244. [Google Scholar]
- Pan, H.; Li, Y.; Chen, L.; Li, J. Molecular processes of dodder haustorium formation on host plant under low Red/Far red (R/FR) irradiation. Int. J. Mol. Sci. 2022, 23, 7528. [Google Scholar] [CrossRef] [PubMed]
- Polli, A.; Souza, L.A.d.; Almeida, O.J.G.d. Structural development of the fruits and seeds in three mistletoe species of Phoradendron (Visceae: Santalaceae). Rodriguésia 2016, 67, 649–659. [Google Scholar] [CrossRef]
- González, F.; Pabón-Mora, N. On embryo polarity, polycotyly, and the homology test of conjunction: A reply to Kuijt´ s (2017) critique. Brittonia 2018, 70, 150–154. [Google Scholar] [CrossRef]
- Kuang, J.; Wang, Y.; Mao, K.; Milne, R.; Wang, M.; Miao, N. Transcriptome Profiling of a Common Mistletoe Species Parasitizing Four Typical Host Species in Urban Southwest China. Genes 2022, 13, 1173. [Google Scholar] [CrossRef]
- Kuijt, J. The anatomy of haustoria and related organs of Gaiadendron (Loranthaceae). Can. J. Bot. 1965, 43, 687–694. [Google Scholar] [CrossRef]
- Sibinelli, V.; Ceccantini, G. Axial and radial wood maturation in three mistletoe ‘giants’(Loranthaceae). IAWA J. 2022, 43, 1–19. [Google Scholar] [CrossRef]
- Calvin, C.L. Anatomy of the endophytic system of the mistletoe, Phoradendron flavescens. Bot. Gaz. 1967, 128, 117–137. [Google Scholar] [CrossRef]
- Mylo, M.D.; Hofmann, M.; Delp, A.; Scholz, R.; Walther, F.; Speck, T.; Speck, O. Advances on the visualization of the internal structures of the European mistletoe: 3D reconstruction using microtomography. Front. Plant Sci. 2021, 12, 715711. [Google Scholar] [CrossRef]
- Natesan, B.; Suvaathimani, S.; Vinothini, K.; Barathan, N. Morphological mimicking and defense mechanism between the hosts and mistletoe: Dendrophthoe falcata of Loranthaceae. Species 2022, 23, 554–565. [Google Scholar]
- Mylo, M.D.; Hofmann, M.; Balle, F.; Beisel, S.; Speck, T.; Speck, O. Biomechanics of the parasite–host interaction of the European mistletoe. J. Exp. Bot. 2022, 73, 1204–1221. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Black, M. Seeds. In Seeds; Springer: Berlin/Heidelberg, Germany, 1994; pp. 1–33. [Google Scholar]
- Ladley, J.J.; Kelly, D. Dispersal, germination and survival of New Zealand mistletoes (Loranthaceae): Dependence on birds. N. Z. J. Ecol. 1996, 20, 69–79. [Google Scholar]
- Ramírez, M.M.; Ornelas, J.F. Germination of Psittacanthus schiedeanus (mistletoe) seeds after passage through the gut of Cedar Waxwings and Grey Silky-flycatchers. J. Torrey Bot. Soc. 2009, 136, 322–331. [Google Scholar] [CrossRef]
- Mellado, A.; Zamora, R. Generalist birds govern the seed dispersal of a parasitic plant with strong recruitment constraints. Oecologia 2014, 176, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.J.; Shamoun, S.F.; Punja, Z.K. In vitro germination and development of western hemlock dwarf mistletoe. Plant Cell Tissue Organ Cult. 2001, 66, 97–105. [Google Scholar] [CrossRef]
- Yoshida, S.; Cui, S.; Ichihashi, Y.; Shirasu, K. The haustorium, a specialized invasive organ in parasitic plants. Annu. Rev. Plant Biol. 2016, 67, 643–667. [Google Scholar] [CrossRef] [Green Version]
- Wickramasuriya, A.; Hewavithana, T.M.; de Silva, K.K.; Ullah, I.; Dunwell, J.M. Comprehensive analysis of Arabidopsis thaliana DNA polymerase epsilon catalytic subunit A and B mutants–an insight into differentially expressed genes and protein-protein interactions. bioRxiv 2022. [Google Scholar] [CrossRef]
- Kodama, Y.; Suetsugu, N.; Kong, S.-G.; Wada, M. Two interacting coiled-coil proteins, WEB1 and PMI2, maintain the chloroplast photorelocation movement velocity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 19591–19596. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, P.a. Structure and evolution of plant disease resistance genes. J. Appl. Genet. 2002, 43, 403–414. [Google Scholar]
- Suh, H.Y.; Kim, J.H.; Woo, J.S.; Ku, B.; Shin, E.J.; Yun, Y.; Oh, B.H. Crystal structure of DeSI-1, a novel deSUMOylase belonging to a putative isopeptidase superfamily. Proteins Struct. Funct. Bioinform. 2012, 80, 2099–2104. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.C.; Olivares, F.L.; Peres, L.E.P.; Piccolo, A.; Canellas, L.P. Plant hormone crosstalk mediated by humic acids. Chem. Biol. Technol. Agric. 2022, 9, 1–25. [Google Scholar] [CrossRef]
- Kaur, M.; Tak, Y.; Bhatia, S.; Asthir, B.; Lorenzo, J.M.; Amarowicz, R. Crosstalk during the carbon–nitrogen cycle that interlinks the biosynthesis, mobilization and accumulation of seed storage reserves. Int. J. Mol. Sci. 2021, 22, 12032. [Google Scholar] [CrossRef] [PubMed]
- Orona-Tamayo, D.; Wielsch, N.; Escalante-Pérez, M.; Svatos, A.; Molina-Torres, J.; Muck, A.; Ramirez-Chávez, E.; Ádame-Alvarez, R.-M.; Heil, M. Short-term proteomic dynamics reveal metabolic factory for active extrafloral nectar secretion by Acacia cornigera ant-plants. Plant J. 2013, 73, 546–554. [Google Scholar] [CrossRef]
- Arc, E.; Galland, M.; Cueff, G.; Godin, B.; Lounifi, I.; Job, D.; Rajjou, L. Reboot the system thanks to protein post-translational modifications and proteome diversity: How quiescent seeds restart their metabolism to prepare seedling establishment. Proteomics 2011, 11, 1606–1618. [Google Scholar] [CrossRef]
- Müntz, K.; Belozersky, M.; Dunaevsky, Y.; Schlereth, A.; Tiedemann, J. Stored proteinases and the initiation of storage protein mobilization in seeds during germination and seedling growth. J. Exp. Bot. 2001, 52, 1741–1752. [Google Scholar] [CrossRef] [Green Version]
- Matilla, A.J. Exploring Breakthroughs in Three Traits Belonging to Seed Life. Plants 2022, 11, 490. [Google Scholar] [CrossRef]
- Zou, Z.; Zhao, Y.; Zhang, L.; Xiao, Y.; Guo, A. Analysis of Cyperus esculentus SMP family genes reveals lineage-specific evolution and seed desiccation-like transcript accumulation during tuber maturation. Ind. Crops Prod. 2022, 187, 115382. [Google Scholar] [CrossRef]
- Pan, L.; Wan, L.; He, L.; Song, L.; Long, H.; Ji, X.; Jiang, N.; Huo, J.; Wei, S.; Fu, J. Comparative proteomic analysis of parasitic loranthus seeds exposed to dehydration stress. Plant Biotechnol. Rep. 2021, 15, 95–108. [Google Scholar] [CrossRef]
- Walters, C.; Berjak, P.; Pammenter, N.; Kennedy, K.; Raven, P. Preservation of recalcitrant seeds. Science 2013, 339, 915–916. [Google Scholar] [CrossRef]
- Wenjing, W.; Chen, Q.; Singh, P.K.; Huang, Y.; Pei, D. CRISPR/Cas9 edited HSFA6a and HSFA6b of Arabidopsis thaliana offers ABA and osmotic stress insensitivity by modulation of ROS homeostasis. Plant Signal. Behav. 2020, 15, 1816321. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Niu, C.-Y.; Yang, C.-R.; Jinn, T.-L. The heat stress factor HSFA6b connects ABA signaling and ABA-mediated heat responses. Plant Physiol. 2016, 172, 1182–1199. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.M.; Del Pozo, J.C.; Walker, L.; Hobbie, L.; Risseeuw, E.; Banks, T.; Crosby, W.L.; Yang, M.; Ma, H.; Estelle, M. Identification of an SCF ubiquitin–ligase complex required for auxin response in Arabidopsis thaliana. Genes Dev. 1999, 13, 1678–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, C.C.S.; Huang, W.N.; Huang, Y.C.; Yeh, K.C. Proteomic survey of copper-binding proteins in Arabidopsis roots by immobilized metal affinity chromatography and mass spectrometry. Proteomics 2006, 6, 2746–2758. [Google Scholar] [CrossRef] [PubMed]
- Holding, D.R.; Otegui, M.S.; Li, B.; Meeley, R.B.; Dam, T.; Hunter, B.G.; Jung, R.; Larkins, B.A. The maize floury1 gene encodes a novel endoplasmic reticulum protein involved in zein protein body formation. Plant Cell 2007, 19, 2569–2582. [Google Scholar] [CrossRef] [Green Version]
- Domon, C.; Lorkovic, Z.J.; Valcárcel, J.; Filipowicz, W. Multiple forms of the U2 small nuclear ribonucleoprotein auxiliary factor U2AF subunits expressed in higher plants. J. Biol. Chem. 1998, 273, 34603–34610. [Google Scholar] [CrossRef] [Green Version]
- Mahalingam, R.; Walling, J.G. Genomic survey of RNA recognition motif (RRM) containing RNA binding proteins from barley (Hordeum vulgare ssp. vulgare). Genomics 2020, 112, 1829–1839. [Google Scholar] [CrossRef]
- Quiroz-Figueroa, F.; Vázquez-Ramos, J.M. Expression of maize D-type cyclins: Comparison, regulation by phytohormones during seed germination and description of a new D cyclin. Physiol. Plant. 2006, 128, 556–568. [Google Scholar] [CrossRef]
- Garza-Aguilar, S.M.; Lara-Núñez, A.; García-Ramírez, E.; Vázquez-Ramos, J.M. Modulation of CycD3; 1-CDK complexes by phytohormones and sucrose during maize germination. Physiol. Plant. 2017, 160, 84–97. [Google Scholar] [CrossRef]
- Masubelele, N.H.; Dewitte, W.; Menges, M.; Maughan, S.; Collins, C.; Huntley, R.; Nieuwland, J.; Scofield, S.; Murray, J.A. D-type cyclins activate division in the root apex to promote seed germination in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 15694–15699. [Google Scholar] [CrossRef] [Green Version]
- Barrôco, R.M.; Van Poucke, K.; Bergervoet, J.H.; De Veylder, L.; Groot, S.P.; Inzé, D.; Engler, G. The role of the cell cycle machinery in resumption of postembryonic development. Plant Physiol. 2005, 137, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Yoshinaga, M.; Inagaki, Y. Ubiquity and origins of structural maintenance of chromosomes (SMC) proteins in eukaryotes. Genome Biol. Evol. 2021, 13, evab256. [Google Scholar] [CrossRef] [PubMed]
- Piao, L.; Yuan, X.; Wang, L.; Xu, X.; Zhuang, M.; Li, J.; Kong, R.; Liu, Z. Loss of histone H4 lysine 20 trimethylation in osteosarcoma is associated with aberrant expression ofhistone methyltransferase SUV420H2. Oncol. Lett. 2020, 20, 26. [Google Scholar] [CrossRef] [PubMed]
- González-Thuillier, I.; Venegas-Calerón, M.; Moreno-Pérez, A.J.; Salas, J.J.; Garcés, R.; von Wettstein-Knowles, P.; Martínez-Force, E. Sunflower (Helianthus annuus) fatty acid synthase complex: β-Ketoacyl-[acyl carrier protein] reductase genes. Plant Physiol. Biochem. 2021, 166, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Chen, K.; Dong, N.Q.; Ye, W.W.; Shan, J.X.; Lin, H.X. Tillering and small grain 1 dominates the tryptophan aminotransferase family required for local auxin biosynthesis in rice. J. Integr. Plant Biol. 2020, 62, 581–600. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ma, B.; Tao, J.-J.; Yin, C.-C.; Hu, Y.; Huang, Y.-H.; Wei, W.; Xin, P.-Y.; Chu, J.-F.; Zhang, W.-K. Rice EIL1 interacts with OsIAAs to regulate auxin biosynthesis mediated by the tryptophan aminotransferase MHZ10/OsTAR2 during root ethylene responses. Plant Cell 2022, 34, 4366–4387. [Google Scholar] [CrossRef]
- Jhu, M.-Y.; Farhi, M.; Wang, L.; Zumstein, K.; Sinha, N.R. Investigating host and parasitic plant interaction by tissue-specific gene analyses on tomato and cuscuta campestris interface at three haustorial developmental stages. Front. Plant Sci. 2022, 12, 3402. [Google Scholar] [CrossRef]
- Gibson, D.M.; King, B.C.; Hayes, M.L.; Bergstrom, G.C. Plant pathogens as a source of diverse enzymes for lignocellulose digestion. Curr. Opin. Microbiol. 2011, 14, 264–270. [Google Scholar] [CrossRef]
- Kantharaj, P.; Boobalan, B.; Sooriamuthu, S.; Mani, R. Lignocellulose degrading enzymes from fungi and their industrial applications. Int. J. Curr. Res. Rev. 2017, 9, 1–12. [Google Scholar]
- Guo, H.; He, T.; Lee, D.-J. Contemporary proteomic research on lignocellulosic enzymes and enzymolysis: A review. Bioresour. Technol. 2022, 344, 126263. [Google Scholar] [CrossRef]
- Silva, J.P.; Ticona, A.R.; Hamann, P.R.; Quirino, B.F.; Noronha, E.F. Deconstruction of lignin: From enzymes to microorganisms. Molecules 2021, 26, 2299. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Dinneny, J.R. A wall with integrity: Surveillance and maintenance of the plant cell wall under stress. New Phytol. 2020, 225, 1428–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafiei, V.; Vélëz, H.; Tzelepis, G. The role of glycoside hydrolases in phytopathogenic fungi and oomycetes virulence. Int. J. Mol. Sci. 2021, 22, 9359. [Google Scholar] [CrossRef] [PubMed]
- Delavault, P.; Montiel, G.; Brun, G.; Pouvreau, J.-B.; Thoiron, S.; Simier, P. Communication between host plants and parasitic plants. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2017; Volume 82, pp. 55–82. [Google Scholar]
- Yoshida, S.; Shirasu, K. Plants that attack plants: Molecular elucidation of plant parasitism. Curr. Opin. Plant Biol. 2012, 15, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Kurotani, K.-i.; Wakatake, T.; Ichihashi, Y.; Okayasu, K.; Sawai, Y.; Ogawa, S.; Cui, S.; Suzuki, T.; Shirasu, K.; Notaguchi, M. Host-parasite tissue adhesion by a secreted type of β-1, 4-glucanase in the parasitic plant Phtheirospermum japonicum. Commun. Biol. 2020, 3, 407. [Google Scholar] [CrossRef]
- Srivastava, M.; Sadanandom, A. An insight into the factors influencing specificity of the sumo system in plants. Plants 2020, 9, 1788. [Google Scholar] [CrossRef]
- Nun, N.B.; Mayer, A.M. Culture of pectin methylesterase and polyphenoloxidasein Cuscuta campestris. Phytochemistry 1999, 50, 719–727. [Google Scholar] [CrossRef]
- Nun, N.B.; Mor, A.; Mayer, A.M. A cofactor requirement for polygalacturonase from Cuscuta campestris. Phytochemistry 1999, 52, 1217–1221. [Google Scholar]
- Bleischwitz, M.; Albert, M.; Fuchsbauer, H.-L.; Kaldenhoff, R. Significance of Cuscutain, a cysteine protease from Cuscuta reflexa, in host-parasite interactions. BMC Plant Biol. 2010, 10, 227. [Google Scholar] [CrossRef]
- Shimizu, K.; Aoki, K. Development of parasitic organs of a stem holoparasitic plant in genus Cuscuta. Front. Plant Sci. 2019, 10, 1435. [Google Scholar] [CrossRef] [Green Version]
- Nagar, R.; Singh, M.; Sanwal, G. Cell wall degrading enzymes in Cuscuta reflexa and its hosts. J. Exp. Bot. 1984, 35, 1104–1112. [Google Scholar] [CrossRef]
- Mayer, A. Pathogenesis by fungi and by parasitic plants: Similarities and differences. Phytoparasitica 2006, 34, 3–16. [Google Scholar]
- Jhu, M.-Y.; Ichihashi, Y.; Farhi, M.; Wong, C.; Sinha, N.R. LATERAL ORGAN BOUNDARIES DOMAIN 25 functions as a key regulator of haustorium development in dodders. Plant Physiol. 2021, 186, 2093–2110. [Google Scholar] [PubMed]
- Nguyen, S.T.; Freund, H.L.; Kasanjian, J.; Berlemont, R. Function, distribution, and annotation of characterized cellulases, xylanases, and chitinases from CAZy. Appl. Microbiol. Biotechnol. 2018, 102, 1629–1637. [Google Scholar] [CrossRef] [PubMed]
- Singhania, R.R.; Patel, A.K.; Sukumaran, R.K.; Larroche, C.; Pandey, A. Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresour. Technol. 2013, 127, 500–507. [Google Scholar]
- Fernández-Sanromán, Á.; Sanromán, M.Á. Extremozymes in food production and processing. In Value-Addition in Food Products and Processing through Enzyme Technology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 25–43. [Google Scholar]
- Queijeiro-Bolaños, M.E.; Malda-Barrera, G.X.; Carrillo-Angeles, I.G.; Suzán-Azpiri, H. Contrasting gas exchange effects on the interactions of two mistletoe species and their host Acacia schaffneri. J. Arid Environ. 2020, 173, 104041. [Google Scholar] [CrossRef]
- Ranjan, A.; Ichihashi, Y.; Farhi, M.; Zumstein, K.; Townsley, B.; David-Schwartz, R.; Sinha, N.R. De novo assembly and characterization of the transcriptome of the parasitic weed dodder identifies genes associated with plant parasitism. Plant Physiol. 2014, 166, 1186–1199. [Google Scholar]
- Cocoletzi, E.; Angeles, G.; Briones, O.; Ceccantini, G.; Ornelas, J.F. The ecophysiology of a neotropical mistletoe depends on the leaf phenology of its tree hosts. Am. J. Bot. 2020, 107, 1225–1237. [Google Scholar]
- Ozturk, M.; Coskuner, K.A.; Serdar, B.; Atar, F.; Bilgili, E. Impact of white mistletoe (Viscum album ssp. abietis) infection severity on morphology, anatomy and photosynthetic pigment content of the needles of cilicican fir (Abies cilicica). Flora 2022, 294, 152135. [Google Scholar] [CrossRef]
- Kleszken, E.; Purcarea, C.; Pallag, A.; Ranga, F.; Memete, A.R.; Miere, F.; Vicas, S.I. Phytochemical Profile and Antioxidant Capacity of Viscum album L. Subsp. album and Effects on Its Host Trees. Plants 2022, 11, 3021. [Google Scholar]
- Kubov, M.; Fleischer, P., Jr.; Rozkošný, J.; Kurjak, D.; Konôpková, A.; Galko, J.; Húdoková, H.; Lalík, M.; Rell, S.; Pittner, J. Drought or severe drought? hemiparasitic yellow mistletoe (Loranthus europaeus) amplifies drought stress in sessile oak trees (Quercus petraea) by altering water status and physiological responses. Water 2020, 12, 2985. [Google Scholar] [CrossRef]
- Hartig, K.; Beck, E. Crosstalk between auxin, cytokinins, and sugars in the plant cell cycle. Plant Biol. 2006, 8, 389–396. [Google Scholar] [PubMed]
- Pennings, S.C.; Callaway, R.M. Parasitic plants: Parallels and contrasts with herbivores. Oecologia 2002, 131, 479–489. [Google Scholar] [CrossRef]
- Kaga, Y.; Yokoyama, R.; Sano, R.; Ohtani, M.; Demura, T.; Kuroha, T.; Shinohara, N.; Nishitani, K. Interspecific signaling between the parasitic plant and the host plants regulate xylem vessel cell differentiation in haustoria of Cuscuta campestris. Front. Plant Sci. 2020, 11, 193. [Google Scholar]
- Douglas Grubb, C.; Zipp, B.J.; Ludwig-Müller, J.; Masuno, M.N.; Molinski, T.F.; Abel, S. Arabidopsis glucosyltransferase UGT74B1 functions in glucosinolate biosynthesis and auxin homeostasis. Plant J. 2004, 40, 893–908. [Google Scholar] [CrossRef]
- de Figueiredo, M.R.A.; Strader, L.C. Intrinsic and extrinsic regulators of Aux/IAA protein degradation dynamics. Trends Biochem. Sci. 2022, 47, 865–874. [Google Scholar] [CrossRef]
- O’Neill, D.P.; Davidson, S.E.; Clarke, V.C.; Yamauchi, Y.; Yamaguchi, S.; Kamiya, Y.; Reid, J.B.; Ross, J.J. Regulation of the gibberellin pathway by auxin and DELLA proteins. Planta 2010, 232, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Tomilov, A.A.; Tomilova, N.B.; Abdallah, I.; Yoder, J.I. Localized hormone fluxes and early haustorium development in the hemiparasitic plant Triphysaria versicolor. Plant Physiol. 2005, 138, 1469–1480. [Google Scholar]
- Pan, L.; Wan, L.; Song, L.; He, L.; Jiang, N.; Long, H.; Huo, J.; Ji, X.; Hu, F.; Fu, J. Comparative Proteomic Analysis Provides New Insights into the Development of Haustorium in Taxillus chinensis (DC.) Danser. BioMed Res. Int. 2022, 2022, 1–9. [Google Scholar] [CrossRef]
- Löffler, C.; Czygan, F.-C.; Proksch, P. Role of indole-3-acetic acid in the interaction of the phanerogamic parasite Cuscuta and host plants. Plant Biol. 1999, 1, 613–617. [Google Scholar] [CrossRef]
- Goyet, V.; Wada, S.; Cui, S.; Wakatake, T.; Shirasu, K.; Montiel, G.; Simier, P.; Yoshida, S. Haustorium inducing factors for parasitic Orobanchaceae. Front. Plant Sci. 2019, 10, 1056. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; da Silva, J.A.T.; Duan, J.; Deng, R.; Xu, X.; Ma, G. Endogenous hormone levels and anatomical characters of haustoria in Santalum album L. seedlings before and after attachment to the host. J. Plant Physiol. 2012, 169, 859–866. [Google Scholar] [CrossRef]
- Mizukami, Y.; Fischer, R.L. Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.-i.; Arai, K.; Aoi, Y.; Tanaka, Y.; Hira, H.; Guo, R.; Hu, Y.; Ge, C.; Zhao, Y.; Kasahara, H. The main oxidative inactivation pathway of the plant hormone auxin. Nat. Commun. 2021, 12, 6752. [Google Scholar]
- Nardozza, S.; Cooney, J.; Boldingh, H.L.; Hewitt, K.G.; Trower, T.; Jones, D.; Thrimawithana, A.H.; Allan, A.C.; Richardson, A.C. Phytohormone and transcriptomic analysis reveals endogenous cytokinins affect kiwifruit growth under restricted carbon supply. Metabolites 2020, 10, 23. [Google Scholar]
- Orona-Tamayo, D.; Wielsch, N.; Blanco-Labra, A.; Svatos, A.; Farías-Rodríguez, R.; Heil, M. Exclusive rewards in mutualisms: Ant proteases and plant protease inhibitors create a lock-key system to protect Acacia food bodies from exploitation. Mol. Ecol. 2013, 22, 4087–4100. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Rivas-Castillo, A.; Orona-Tamayo, D.; Gómez-Ramírez, M.; Rojas-Avelizapa, N.G. Diverse molecular resistance mechanisms of Bacillus megaterium during metal removal present in a spent catalyst. Biotechnol. Bioprocess Eng. 2017, 22, 296–307. [Google Scholar] [CrossRef]
- Huynh, M.-L.; Russell, P.; Walsh, B. Tryptic digestion of in-gel proteins for mass spectrometry analysis. In Two-Dimensional Electrophoresis Protocols; Springer: Berlin/Heidelberg, Germany, 2009; pp. 507–513. [Google Scholar]
- Shevchenko, A.; Sunyaev, S.; Loboda, A.; Shevchenko, A.; Bork, P.; Ens, W.; Standing, K.G. Charting the proteomes of organisms with unsequenced genomes by MALDI-quadrupole time-of-flight mass spectrometry and BLAST homology searching. Anal. Chem. 2001, 73, 1917–1926. [Google Scholar] [CrossRef]
- Xiao, Z.; Storms, R.; Tsang, A. Microplate-based carboxymethylcellulose assay for endoglucanase activity. Anal. Biochem. 2005, 342, 176–178. [Google Scholar] [CrossRef]
- Saha, B.C. Production, purification and properties of xylanase from a newly isolated Fusarium proliferatum. Process Biochem. 2002, 37, 1279–1284. [Google Scholar] [CrossRef]
- Zhang, H.; Xie, X.; Kim, M.S.; Kornyeyev, D.A.; Holaday, S.; Paré, P.W. Soil bacteria augment Arabidopsis photosynthesis by decreasing glucose sensing and abscisic acid levels in planta. Plant J. 2008, 56, 264–273. [Google Scholar] [CrossRef]
- Uc-Chuc, M.A.; Pérez-Hernández, C.; Galaz-Ávalos, R.M.; Brito-Argaez, L.; Aguilar-Hernández, V.; Loyola-Vargas, V.M. YUCCA-mediated biosynthesis of the auxin IAA is required during the somatic embryogenic induction process in Coffea canephora. Int. J. Mol. Sci. 2020, 21, 4751. [Google Scholar] [CrossRef]
- Wang, X.-Q.; Pan, X.-J.; Lin, G.-Y.; Xiang, Z.; Wang, X.-B.; Wu, J.-Z.; Lin, D. Simultaneous determination of clozapine, olanzapine and mirtazapine in human plasma by LC-MS/MS. Fa Yi Xue Za Zhi 2009, 25, 123–126. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | Description | Accesion | MS BLAST | Organism | Function | Mw | pI |
|---|---|---|---|---|---|---|---|
| Score | kDa | ||||||
| Gene regulation | |||||||
| 1 | RNA-binding protein 1-like isoform X2 | XP_010322224.1 | 412 | Solanum lycopersicum | Translation, ribosomal structure and biogenesis | 22.2 | 4.4 |
| 5 | U2 small nuclear ribonucleoprotein A′ | XP_004235867.1 | 491 | Solanum lycopersicum | Nuclear mRNA splicing | 32.3 | 5.5 |
| 6 | DNA ligase 1-like | XP_009765758.1 | 297 | Nicotiana sylvestris | DNA recombination and DNA repair | 55.0 | 5.0 |
| 9 | Heat shock transcription factor A6B | NP_188922.1 | 759 | Arabidopsis thaliana | DNA binding | 46.7 | 5.0 |
| 13 | Protein N-lysine methyltransferase METTL21A | XP_004244751.1 | 549 | Solanum lypersicum | Methyltransferase of heat shock protein 70 (HSP70) family members | 31.7 | 4.9 |
| 14 | RNA-binding ASCH domain protein | NP_001324874.1 | 449 | Arabidopsis thaliana | RNA-binding domain during coactivation | 27.4 | 5.2 |
| 23 | Evolutionarily conserved C-terminal region 9 | NP_174117.2 | 1077 | Arabidopsis thaliana | RNA binding | 61.2 | 6.3 |
| 25 | AP2-like ethylene-responsive transcription factor AIL1 | NM_001366759.1 | 1080 | Solanum lypersicum | DNA binding | 62.0 | 6.0 |
| 29 | DNA replication licensing factor MCM2 | XP_004250699.2 | 1854 | Solanum lycopersicum | 3′-5′ DNA helicase/THO complex | 108.1 | 5.1 |
| 31 | 5′-3′ exonuclease family protein | NP_001325748.1 | 2779 | Arabidopsis thaliana | Exonuclease that digests recessed strands of DNA duplexes in the 3′ to 5′ direction | 168.8 | 4.9 |
| Cell division | |||||||
| 2 | Cyclin-D5-3-like | XP_019069003.1 | 316 | Solanum lycopersicum | Cell-cycle | 24.5 | 4.7 |
| 30 | WEB family protein At3g02930, chloroplastic | XP_004235278.1 | 1399 | Solanum lycopersicum | Microtubule motor activity | 109.9 | 4.9 |
| 32 | Leucine-rich repeat-containing protein DDB_G0290503 | XP_010674981.1 | 1019 | Beta vulgaris subsp. vulgaris | Probable component of the transverse filaments | 71.6 | 5.4 |
| Photosynthesis | |||||||
| 27 | Magnesium-chelatase subunit ChlD, chloroplastic | XP_004236627.1 | 1327 | Solanum lypersicum | Chlorophyll biosynthesis | 83.9 | 5.4 |
| Metabolism | |||||||
| 3 | SKP1-like protein 11 | XP_010321926.2 | 368 | Solanum lypersicum | E3 ligase complex | 21.0 | 5.0 |
| 7 | 3-oxoacyl-acyl-carrier synthase-like protein | NP_196072.1 | 148 | Arabidopsis thaliana | Fatty acid synthesis | 32.9 | 6.2 |
| 8 | deSI-like protein At4g17486 isoform X1 | XP_010325508.1 | 401 | Solanum lypersicum | Thiol peptidase | 24.9 | 6.2 |
| 10 | Zein-binding protein | NP_200591.2 | 786 | Arabidopsis thaliana | Zein-binding domain-containing protein | 44.0 | 5.2 |
| 11 | Pectinesterase 15 isoform X2 | XP_048504660.1 | 101 | Beta vulgaris subsp. vulgaris | Acts in the modification of cell walls via demethylesterification of cell wall pectin | 15.6 | 5.9 |
| 12 | Sugar transporter, putative | NP_569015.1 | 261 | Arabidopsis thaliana | Sugar transporter | 18.2 | 5.2 |
| 19 | RING/U-box superfamily protein | NP_196267.1 | 309 | Arabidopsis thaliana | E3 ubiquitin ligase | 22.0 | 6.4 |
| 20 | Chaperone DnaJ-domain superfamily protein | NP_174319.1 | 650 | Arabidopsis thaliana | Folding and degradation of proteins | 50.7 | 5.3 |
| Phytohotmone pathway signaling | |||||||
| 21 | Tryptophan aminotransferase related 2 | NP_567706.1 | 887 | Arabidopsis thaliana | Trytophan aminotransferase/C-S-lyase | 50.0 | 6.1 |
| Development | |||||||
| 18 | Transducin/WD40 repeat-like superfamily protein | NP_851281.1 | 675 | Arabidopsis thaliana | Regulation of dynamic multi-subunit complexes | 36.1 | 6.2 |
| 24 | Phosphatidylinositol 4-kinase gamma 3 | XP_010671446.1 | 1150 | Beta vulgaris subsp. vulgaris | Phosphatidylinositol 3-and 4-kinase | 63.8 | 5.7 |
| 28 | Exocyst complex component SEC5 | XP_010680634.1 | 2079 | Beta vulgaris subsp. vulgaris | Involved in polarized cell growth | 122.8 | 5.6 |
| Cellular response | |||||||
| 4 | PYK10-binding protein 1 | NP_001030710.1 | 530 | Arabidopsis thaliana | Inhibitor-type lectin that may regulate the correct polymerization of BGLU23/PYK10 upon tissue damage | 32.1 | 5.8 |
| 15 | Bacteriophage N4 adsorption B protein | NP_569035.1 | 104 | Arabidopsis thaliana | Immune response | 72.0 | 5.0 |
| 16 | NAC domain-containing protein 74-like | XP_016449206.1 | 296 | Nicotiana tabacum | Transcription activator involved in heat and endoplasmic reticulum (ER) stress responses | 44.4 | 5.3 |
| 17 | 2-oxoglutarate-dependent dioxygenase-like protein | NP_001322930.1 | 653 | Arabidopsis thaliana | Glucosinolates biosynthesis | 37.7 | 6.3 |
| 22 | Protein DJ-1 homolog B-like | XP_010675408.1 | 897 | Beta vulgaris subsp. vulgaris | Glyoxalase I activity | 47.1 | 6.3 |
| 26 | Pleckstrin homology (PH) domain-containing protein | NP_850155.1 | 831 | Arabidopsis thaliana | Binds specifically to phosphatidylinositol 3-phosphate (PtdIns3P) | 56.0 | 5.9 |
| Infective Stages | |||||||
|---|---|---|---|---|---|---|---|
| Phytohormone | Abbreviation | T1 | T2 | T3 | T4 | T5 | Leaf |
| Auxins | |||||||
| Indole-3-Acetic Acid | IAA | 1.3 ± 0.2 c | 0.2 ± 0.01 a | ND | 0.6 ± 0.1 b | 0.3 ± 0.02 a | ND |
| Indole-3-Acetyl-L-Aspartic Acid | IAA-L-Asp | 71.8 ± 10.3 c | 8.0 ± 3.4 ab | 7.0 ± 0.4 b | 3.9 ± 0.1 a | 6.1 ± 0.79 b | 2.9 ± 0.3 a |
| Indole-3-Acetyl-L-Glutamic Acid | IAA-L-Glu | 913.6 ± 170.9 e | 121.9 ± 26.8 d | 90.1 ±5.2 c | 2.1 ± 0.1 b | 2.7 ± 0.3 b | 0.5 ± 0.05 a |
| Indole-3-Acetyl-Alanine | IAA-L-Ala | ND | ND | ND | ND | ND | ND |
| Cytokinins | |||||||
| Dehydrozeatin | DZ | 1.3 ± 0.2 b | 3.2 ± 0.6 c | 0.4 ± 0.03 a | ND | ND | ND |
| N6-furfuryladenine | Kinetin | ND | ND | ND | ND | ND | ND |
| N6-Isopentenyladenine | iP | 0.1 ± 0.01 a | 2.6 ± 0.34 b | 0.2 ± 0.01 a | ND | ND | ND |
| N6-benzyladenine | BA | ND | ND | ND | ND | ND | ND |
| cis-Zeatin-Riboside | cZR | 0.2 ± 0.03 b | 0.9± 0.2 c | 0.2 ± 0.02 b | ND | 0.9 ± 0.15 c | 0.08 ± 0.01 a |
| trans-Zeatin | tZ | ND | 0.5 ± 0.1 a | ND | ND | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar-Venegas, M.; Quintana-Rodríguez, E.; Aguilar-Hernández, V.; López-García, C.M.; Conejo-Dávila, E.; Brito-Argáez, L.; Loyola-Vargas, V.M.; Vega-Arreguín, J.; Orona-Tamayo, D. Protein Profiling of Psittacanthus calyculatus during Mesquite Infection. Plants 2023, 12, 464. https://doi.org/10.3390/plants12030464
Aguilar-Venegas M, Quintana-Rodríguez E, Aguilar-Hernández V, López-García CM, Conejo-Dávila E, Brito-Argáez L, Loyola-Vargas VM, Vega-Arreguín J, Orona-Tamayo D. Protein Profiling of Psittacanthus calyculatus during Mesquite Infection. Plants. 2023; 12(3):464. https://doi.org/10.3390/plants12030464
Chicago/Turabian StyleAguilar-Venegas, Montserrat, Elizabeth Quintana-Rodríguez, Víctor Aguilar-Hernández, Claudia Marina López-García, Efraín Conejo-Dávila, Ligia Brito-Argáez, Víctor M. Loyola-Vargas, Julio Vega-Arreguín, and Domancar Orona-Tamayo. 2023. "Protein Profiling of Psittacanthus calyculatus during Mesquite Infection" Plants 12, no. 3: 464. https://doi.org/10.3390/plants12030464